Après de nombreux travaux qui se sont rapidement succédés sur ce sujet tant en France qu'à l'étranger, la nature du tubercule des Ophrydées est encore en litige. Ayant déjà pris une humble part à la lutte, je m'aventure à redescendre dans la lice avec de nouvelles armes, savoir : la germination des Ophrydées, quelques points remarquables de botanique comparée et des études sur les tubercules palmés. Mes forces trahiront peut-être mon courage en face d' un problème si controversé ; j'ai cependant l'espoir que mes observations ne seront pas complètement inutiles pour la solution qui tarde tant à se manifester, car la germination des Ophrydées a été jusqu'ici peu étudiée, et le fait capital des premières évolutions de la plante me paraît avoir échappé aux observateurs qui m'ont précédé sur ce terrain. C'est du moins ce que je crois pouvoir déduire des notes et croquis calqués sur le travail de Salisbury (A. Salisbury, On the germinat. of the seeds of Orchideoe, in Trans. linn societ., VII), et que je dois à l'obligeance de M. Clos.
§ 1. Germination.
C'est à des circonstances purement fortuites que je dois d'avoir assisté à la germination des Ophrydées. Vers la fin de mars, en arrachant dans les alluvions limoneuses de la Durance un pied d'Ophrys apifera, j'aperçus, sous une mince couche d'humus, un grand nombre de corpuscules bulbiformes disséminés tout autour de la plante dont je convoitais le tubercule. D'autres pieds d'Ophrys furent visités; presque tous, dans le voisinage de leur base, étaient accompagnés des mêmes productions. Soupçonnant déjà la nature de ma découverte, je multipliai mes recherches et j'eus la satisfaction de rencontrer tous les états intermédiaires entre ces corpuscules et de très jeunes plantules d'Ophrys, montrant à l'air la pointe verte de leur première feuille. J'avais donc sous les yeux la germination de l'Ophrys apifera. Je remplis ma boîte d'herborisalion du précieux terreau qui renfermait ces corpuscules pour le soumettre, à loisir, à une patiente exploration avec le secours de la loupe, ce qui me permit, avec les plantules plus on moins avancées dont je fis provision, d'assister en peu de temps à toutes les phases de la végétation de la plante pendant sa première année.
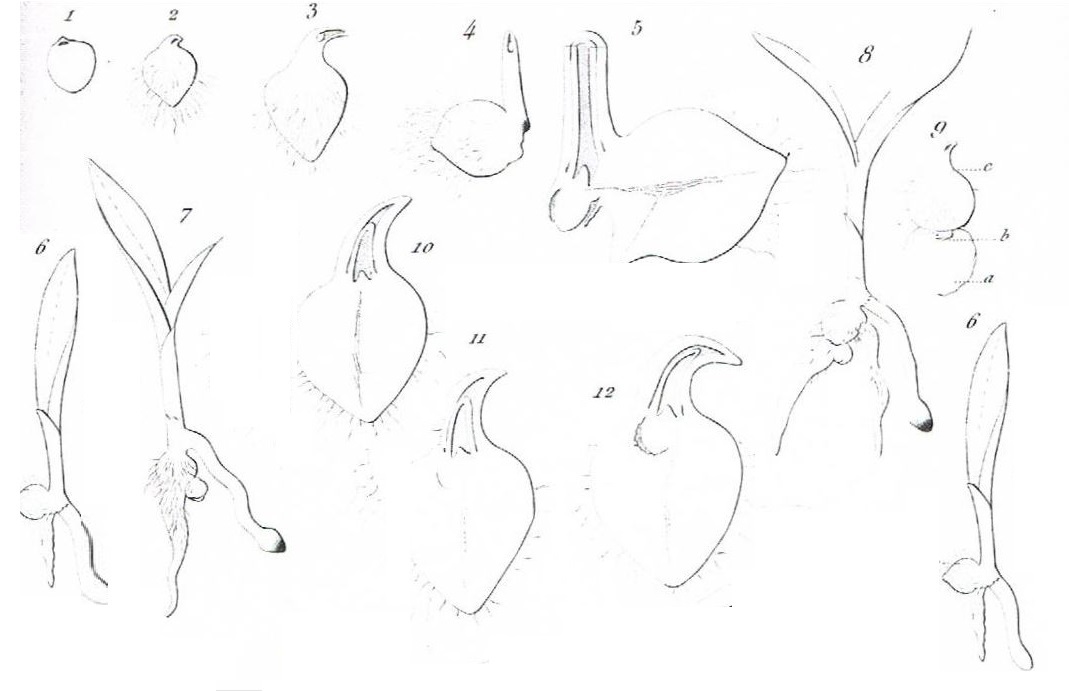
Fig 1,2,3,4,5,6. Germination de l'Ophrys apifera
Fig. 1. La masse embryonnaire quand elle est débarrassée de ses téguments. Diamètre, 1/4 à 1/3 de millimètre.
Fig. 2. La même après l'apparition de la villosité du tubercule.Diamètre, 1 millimètre.
Fig. 3 et 4. La même, à des degrés différents.
Fig. 5. Coupe d'une plantule pereille à celle de la figure 4, montrant l'origine de ses deux mamalons.
Fig. 6. La plantule avec le développement qu'elle doit acquérir la première année. Grandeur naturelle.
Fig 7,8,9. Ophrys apifera dans sa seconde année.
Fig 7,8. La plante seconde année. Grandeur naturelle.
Fig 9. L'une des premières formes sous lesquelles se montre la plante de seconde année. Le renflement inférieur est le tubercule terminal produit la première année. Diamètre de ce tubercule, 1 à 3 millimètre
Fig 10,11,12,13,14. Développement et structure du tubercule terminal produit la première année. Diamètre de ce tubercule, 1 à 3 millimètre
Fig 10,11,12,13. Coupe de plantules pareilles à celles des figures 2,3,4 pour montrer l'organisation graduelle de la gemme terminale en tubercule.
Confiées au sol depuis la dernière floraison, depuis près d'un an, les graines microscopiques de l'Ophrys ont subi comme une longue incubation, pendant laquelle l'embryon, sans changer sensiblement de forme, a acquis un volume plus considérable, en puisant directement dans l'humus et par toute sa surface quelques sucs assimilables. Les graines en petit nombre que la loupe me fait découvrir sont totalement déformées. L'enveloppe externe, lâche et fusiforme dans le principe, est actuellement distendue outre mesure, et crève sous la pression d'un noyau interne, qu'elle ne peut plus contenir. Ce noyau (pl. 11, fig. 1) est un sphéroïde, dont le diamètre mesure de 1/4 à 1/3 de millimètre. Sa surface est lisse, luisante et d'un aspect un peu framboisé, dû aux cellules externes, amples et gonflées. Sa substance est presque aussi limpide que du cristal, si ce n'est vers l'extrémité inférieure où se montre une faible nébulosité, occasionnée probablement par l'apparition des premiers grains de fécule. A l'extrémité opposée se trouve un ostiole à lèvres étroitement contractées ; l'une de ces lèvres s'élève un peu plus que l'autre, et forme un léger bourrelet. Ce corpuscule limpide, c'est l'embryon, dont la forme est encore la même que dans la graine récemment disséminée ( voyez A. de Jussieu, Elem., p 583, fig 556). Débarrassé de ses enveloppes, le globule embryonnaire se couvre, excepté dans sa partie supérieure, d'une épaisse villosité formée de filaments blancs, longs et soyeux. Si l'on met l'embryon dans une goutte d'eau, ces filaments, qui ne sont autre chose que des prolongements pileux des cellules épidermiques, s'étalent, et forment autour de lui une épaisse chevelure aranéeuse. Peut-être jouent-ils un rôle dans la nutrition de ce délicat organisme, qui, isolé, sans radicule, doit pourtant arriver ainsi à un volume assez considérable. On est d'autant plus porté à le croire, qu'on voit ces filaments s'irradier dans tous les sens, s'insinuer dans le sol comme autant de suçoirs se portant au devant des sucs nutritifs. En même temps que cette chevelure apparaît, le corps embryonnaire se façonne un peu en pointe à son pôle inférieur, et de globuleux devient conoïde. Cette extrémité aiguë a une teinte jaunâtre, due en grande partie à une pellicule en lambeaux, dernier débris des téguments de la graine. Au pôle supérieur, des changements notables ont également lieu ; les lèvres de l'ostiole, fortement relevées de toute part, forment un capuchon légèrement recourbé, et muni vers l'extrémité d'une étroite boutonnière. Ce capuchon, c'est la feuille colylédonnaire ; cette fente, l'orifice par où doit sortir, non l'axe de la plante, mais simplement la feuille suivante. L'axe, en effet, au lieu de s'engager dans cette route normale, doit s'ouvrir lui-même un passage, que rien encore ne fait soupçonner. La plantule rudimentaire a alors 1 millimètre environ en longueur (pl. 11, fig. 2).
Sans éprouver d'autres modifications remarquables, le cotylédon continue à s'allonger, tout en s'incurvant toujours davantage ; en même temps, sa fente s'épanouit, et laisse apercevoir un petit mamelon qui est la pointe de la feuille suivante (fig. 3). Enfin son incurvation devient telle, que sa direction est perpendiculaire à l'axe du renflement villeux qui le supporte. Mais alors deux faibles gibbosités ne tardent pas à se dessiner sur la partie convexe de la plantule, presque immédiatement l'une au-dessus de l'autre. L'inférieure appartient au renflement villeux; la supérieure paraît naître de la base du cotylédon. Généralement le mamelon supérieur se montre un peu avant l'autre. Dans cet état, la feuille cotylédonaire atteint 2 ou 3 millimètres en longueur, et le renflement 2 millimètres environ en diamètre. Celui-ci, à cette époque, a pris une teinte jaunâtre, qui se rembrunit de plus en plus avec l'âge. Sous la pression du mamelon supérieur, la couche superficielle de la plantule se déchire irrégulièrement, et laisse émerger un corps singulier, qu'on prendrait pour une racine, si la différence d'aspect et de coloration qu'il présente avec le produit du mamelon inférieur ne faisait naître des doutes. Si l'on fait sur la plantule une section longitudinale passant par les deux mamelons (fig. 5) on voit, en effet que ces deux organes n'ont rien de commun ; le supérieur étant l'extrémité de l'axe organisée en tubercule, et s'ouvrant un passage à travers l'épaisseur des tissus; le second étant simplement une racine. Arrivée à ce point, la plantule montre toutes les parties qui doivent la composer la première année : un cotylédon, une feuille ordinaire, un renflement tubéreux, un tubercule en voie de formation et une racine. Ces parties continuent à croître, à se perfectionner ; la feuille sort de la gaine du cotylédon et se déploie ; la racine s'allonge tout en se couvrant d'une légère villosité ; le tubercule mûrit appendu à un long pédicelle, et l'on obtient enfin une jeune plante, comme celle représentée de grandeur naturelle dans la figure 6. Le tubercule, produit aux dépens de la sommité de l'axe, passe du blanc mat au roussâtre, se couvre du même duvet qui revêt le renflement inférieur, et résorbe à son profit les sucs amassés dans ce dernier, qui graduellement se ride, devient flasque, se dessèche et se réduit finalement à une pellicule. Mais alors feuille, cotylédon, racine, tout est détruit, et il ne reste plus de la plantule que le tubercule terminal, dépositaire d'un foyer de vie qui doit se réveiller l'année suivante. Le diamètre du tubercule ainsi formé mesure de 1 à 3 millimètres. Constatons, avant d'aller plus loin, une étroite ressemblance entre le renflement tubéreux de la base et le tubercule terminal. Nous les avons vus passer tous les deux par les mêmes nuances de coloration, et se couvrir également d'une même villosité aranéeuse ; mais là ne s'arrête pas la ressemblance, la structure infime est la même. Si l'on coupe transversalement une mince rondelle, soit du renflement inférieur, soit du tubercule, on trouve de part et d'autre : 1° un seul faisceau vasculaire occupant l'axe géométrique ; 2° une zone cellulaire d'un beau blanc, gorgée de très petits grains de fécule ; 3° une zone également cellulaire, mais opaline, et dont les cellules contiennent un gros noyau indivis, qui pourrait bien être le premier état d'une agglomération de granules amylacés, ou peut-être encore une pelote de bassorine ; 4° une pellicule épidermique, dont quelques cellules contiennent chacune un paquet de raphides. Cela constaté, et avant de m'occuper de l'interprétation des divers organes que je viens de passer rapidement en revue, je suivrai l'évolution de la plante pendant la seconde année, car nous trouverons là quelques données pour cette interprétation.
Avec les plantules provenant de la germination des graines, je pus recueillir, dans la même localité, d'autres jeunes pousses presque identiques avec les précédentes, mais munies à leur base d'un tubercule en tout pareil à celui que je viens de décrire (fig. 7 et 8). Ces pousses sont évidemment dans leur seconde année. Si, sur ce tubercule, nous superposons successivement par la pensée les renflements embryonnaires que reproduisent les figures 1, 2 et 3, nous aurons exactement les premières évolutions de la plante dans sa seconde période annuelle. La figure 9 se rapporte à l'un de ces états : a est le tubercule formé l'année précédente; b, la cicatrice que l'axe et les premières formations appendiculaires ont laissée sur lui en se détachant; c, la nouvelle pousse. Il est très digne de remarque de trouver une ressemblance aussi parfaite entre le développement de l'embryon et celui de la gemme du tubercule. Si l'on compare le produit c de cette gemme avec la plante embryonnaire (fig. 2 et 3), il est impossible de trouver la moindre différence ; c'est la même forme, le même volume, la même coloration, le même duvet ; et si le tubercule n'était pas là pour attester leur origine différente, on confondrait inévitablement les deux productions. On doit s'attendre d'après la similitude complète que le bourgeon tubéreux (c, fig. 9) présente avec l'embryon, à retrouver chez lui le même mode d'évolution. La prévision est parfaitement fondée : ainsi la fente du capuchon, formé par la première écaille, livre passage à une seconde écaille, puis à deux feuilles normales, mais ne reçoit pas l'axe qui s'ouvre, comme précédemment, une issue violente, et, organisé à l'extrémité en tubercule, s'enfonce profondément dans le sol (fig. 7 et 8). A la base de la tubérosité inférieure de ces pousses s'allonge, étroitement appliquée contre le vieux tubercule, une racine napiforme, d'un diamètre disproportionné, villeuse comme colle tubérosité, et gonflée comme elle de granules amylacés (fig. 7). Plus tard, mais non toujours, une seconde racine, sans caractères particuliers, apparaît sur le flanc du même renflement, au-dessous du point où se fait jour, précisément comme cela a lieu dans la plantule directement issue de la graine ; de manière qu'en enlevant à la pousse de seconde année (fig. 8) le vieux tubercule et la racine napiforme, on ne pourrait plus la distinguer de celle de première année (fig. 6).
On peut maintenant se rendre compte de la structure de la jeune plante venue de germination. La tubérosité qui lui sert de base se retrouve dans les pousses de seconde année, avec les caractères les plus manifestes d'une portion d'axe d'un premier entre-noeud. Cette tubérosité est donc une tigelle, et l'expression que Salisbury emploie au sujet de l'Orchis Morio, radicula mox in tuber intu mescens (Loc. cit., p. 34.), n'est nullement fondée. Mais où est alors la radicule ? Faut-il regarder comme telle la racine qui s'échappe du flanc de la tigelle au-dessous du point où l'axe se fait jour ? Je ne le crois pas, parce que cette racine est trop volumineuse ; qu'elle n'occupe pas la place qu'elle devrait occuper dans celle hypothèse, c'est -à-dire le pôle inférieur de la masse tigellaire ; parce qu'elle persiste pendant toute la première période annuelle de la plante, et enfin parce que dans les pousses de seconde année une racine pareille se montre exactement à la même place. Or, dans ce dernier cas, c'est évidemment une racine adventive ; pourquoi pas alors dans le premier ? L'embryon de l'Ophrys apifera est donc sans radicule, ce qui n 'est pas d'ailleurs sans exemple dans d'autres plantes, et le premier tubercule produit est le premier entre-noeud, la tigelle, le collet, pour me servir de la nomenclature adoptée par M. Clos, qui a déjà cherché à déterminer la nature de ce premier tubercule ( M. Clos, Du collet dans les plantes, etc., Ann. des sc. nat., 3e série, t. XIII). Appelons, pour éviter des circonlocutions embarrassante, ce premier tubercule tubercule tigellaire ; celui qui se développe au sommet de l'axe, tubercule terminal ; et enfin tubercule basilaire, celui qui s'organise à la base du même axe la seconde année, et même les années suivantes. Il y a identité presque complète entre le tubercule tigellaire et le tubercule basilaire ; la seule différence consiste dans la présence chez ce dernier de la racine napiforme, qui ne se montre jamais dans le premier. Il y a d'autre part, si l'on se rappelle la comparaison que j'en ai donnée plus haut, ressemblance aussi complète que possible, tant à l'extérieur qu'à l'intérieur, entre le tubercule tigellaire et le premier tubercule terminal. La différence porte encore sur une racine adventive qui se développe sur le tubercule tigellaire, et n'apparaît jamais dans l'autre. Mais une différence analogue, quoique plus forte, ne nous empêche pas de proclamer la nature commune des deux premiers tubercules comparés ; pourquoi nous en empêcherait-elle quand nous comparons les deux derniers ? D'ailleurs ces diverses racines adventives sont des caractères purement spécifiques ; car, d'après les figures qu'en a données Salisbury, le tubercule tigellaire de l'Orchis Morio en est complètement dépourvu. Je crois donc me conformer à une rigoureuse logique en regardant les trois tubercules comme des produits analogues, en les considérant tous les trois comme des entre-noeuds ou des portions d'entre-noeud où s'amasse un dépôt féculent. Ainsi l'axe de l'Ophrys apifera se renfle de distance en distance à plusieurs reprises, et peut présenter simultanément jusqu'à trois tubercules superposés (fig. 8) ; savoir : un tubercule inférieur appartenant aux produits de l'année précédente, puis un tubercule intermédiaire plein de vigueur, et enfin un tubercule terminal en voie de formation. N'est-ce pas là l'agencement des renflements bulbiformes formés par les entre-noeuds intérieurs d'un chaume d'Avena bulbosa, renflements disposés bout à bout comme des grains de chapelet ? Pour qu'il y ait parité entre les deux organisations, ce n'est plus qu'une affaire de temps, Que l'Ophrys, au lieu de produire ses tubercules de loin en loin, d'année en année, les produise tous la première année, sans intervalle de repos; que son axe, après ces entre-noeuds hypertrophiés, s'allonge en d'autres entre-noeuds normaux pour se terminer enfin par un épi floral ou par une lige stérile, et nous aurons, dans ce qu'elle présente d'essentiel, la structure d'une tige d'Avena bulbosa. J'ai supposé, pareillement à ce qui a lieu chez cette dernière plante, que l'axe de l'Ophrys, après un nombre indéterminé d'entre-noeuds renflés en tubercules, en produisait enfin de normaux ; c'est ce qui a lieu en effet. Il arrive une époque, après trois ou quatre ans, autant que je peux en juger, où l'axe, issu de la graine, cesse de métamorphoser son extrémité en tubercule et de se renfler à la base. Alors celle extrémité se développe comme à l'ordinaire, mais en restant stérile, et les bourgeons latéraux sont chargés de produire les pousses florales, après plusieurs générations, par voie de gemmation. En résumé, la structure du chaume de I'Avena bulbosa et celle de la tige primordiale de I'Ophrys apifera présentent une similitude frappante : c'est de part et d'autre une série plus ou moins longue de tubercules conjugués provenant des entre-noeuds inférieurs, et à la suite de ces tubercules une tige normale florifère ou stérile. Dans la première plante, ce chapelet de tubercules se forme dans une même saison, et persiste pour alimenter l'année suivante les bourgeons axillaires qui en dépendent ; dans la seconde, les divers tubercules n'existent pas tous simultanément, mais ils apparaissent à de longues périodes pour se transmettre de l'un à l'autre la gemme terminale, dont ils sont chargés d'assurer la complète évolution.
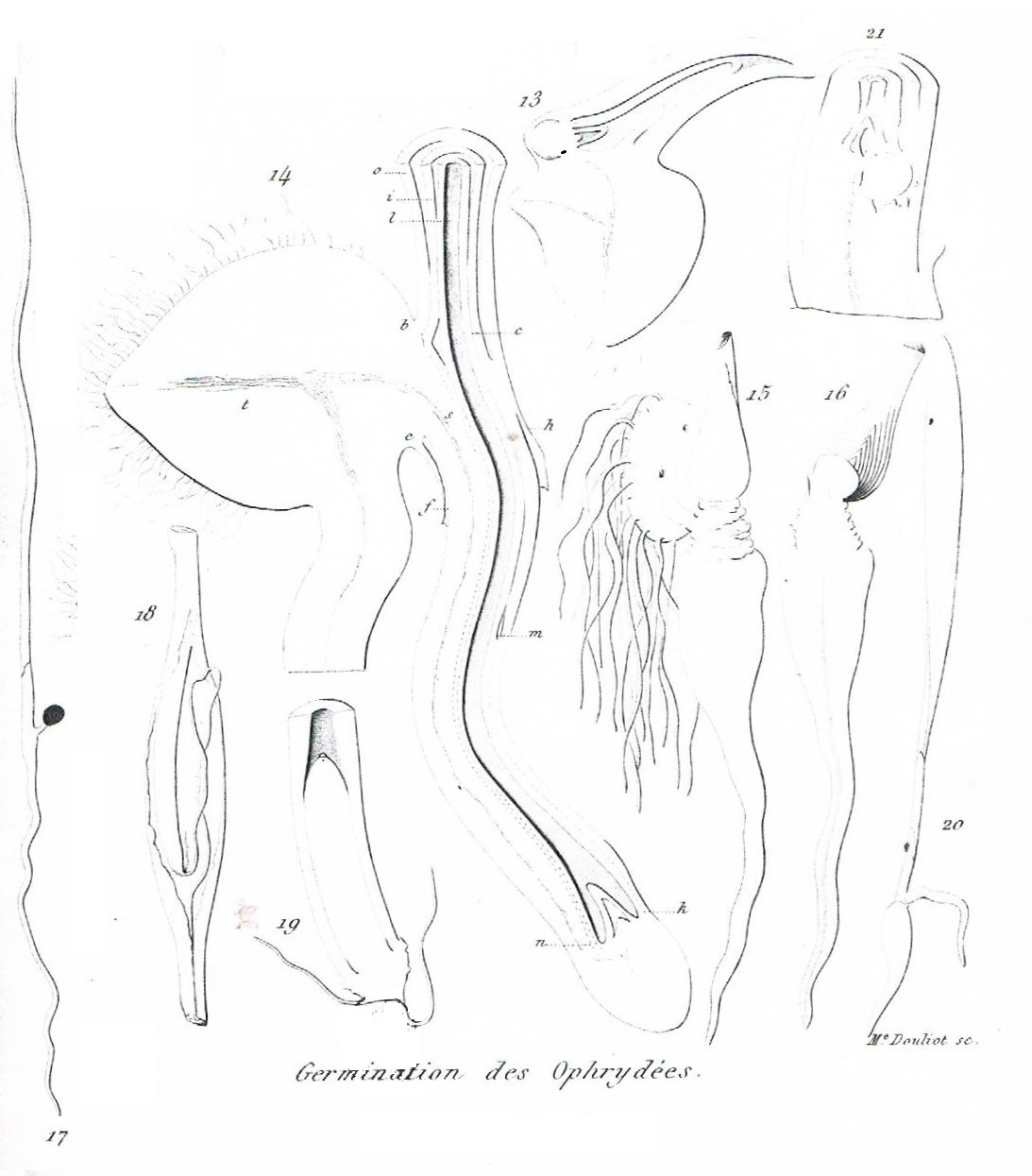
Ophrys apifera dans sa seconde année.
Fig. 14. Coupe de la partie inférieure de la plantule, figure 6, pour montrer la structure du pédicelle du tubercule terminal
Crocus sativus, grandeur naturelle.
Fig. 15. Un des tubercules de Safran dépouillé de ses feuilles, et sur lequel on n'a conservé qu'une pousse de nouvelle formation.
Fig. 16. Coupe de cette pousse, montrant les connexions du tubercule en voie de formation et de sa racine napiforme.
Fig. 17, 18, 19. Germination du Colchicum autumnale.
Fig. 17. La plantule telle qu'elle est avant le déplacement de la gemme. Grandeur naturelle.
Fig. 18. Moitié supérieure de la radicule dans la quelle plonge un cylindre dont l'extréminté inférieure est occupée par la gemme d'abord placée au niveau supérieur de la même figure.
Fig. 19. Coupe de la partie inférieure de ce cylindre, pour montrer le tuberculeterminal qu'il contient ; à sa base sont deux filaments vasculaires, dont l'un est un débris de la nervure du cotylédon, l'autre le cordon vasculaire de la racine.
Fig. 20, 21. Orchis divaricata.
Fig. 20. Plantule d'Orchis divaricata produisant un tubercule terminal. Grandeur naturelle.
Fig. 21. Coupe de la gemme de cette pousse.
La racine adventive qui naît de la partie inférieure du tubercule basilaire mérite un instant notre attention. D'un volume disproportionné avec celui du reste de la plante, elle est, comme le tubercule, roussâtre et couverte d'une villosité touffue ; tantôt elle est nettement séparée du tubercule par un étranglement profond (fig. 8) ; tantôt, au contraire, elle continue le premier d'une manière si insensible, qu'il est impossible, même an moyen d'une loupe, de préciser où le tubercule finit et où la racine commence (fig. 7). On prendrait alors cet ensemble pour un organe simple, si l'on n'était averti par l'examen des plantes où la ligne de démarcation des deux parties est on ne peut plus manifeste. Cette racine est évidemment un réservoir nutritif supplémentaire, concourant au même but que le renflement de la base de la tige, c'est-à-dire à la formation du tubercule terminal. M. Germain de Saint-Pierre a observé des racines analogues chez plusieurs Liliacées bulbeuses (Bull. de la Soc. bot. de France, n° de septembre 1854, p. 166.). J'ai fait de mon côté quelques observations pareilles. Mais le plus bel exemple que je connaisse de ces racines singulières se trouve dans le Safran (Crocus sativus). On sait que dans cette plante, comme dans le Colchique, les entre-noeuds inférieurs de l'axe se renflent en une masse féculente, en un tubercule revêtu par les bases fibreuses et engainante des feuilles. Ce tubercule nourrit de nombreux bourgeons, qui s'organisent à leur tour en tubercules pareils au premier, et sont d'autant plus gros qu'ils partent d'un point plus élevé. Tant que persiste la plante mère, tant que le tubercule. nourricier n'est pas épuisé, les jeunes pousses n'émettent pas de racines. Ce n'est qu'après la résorption complète du tubercule qui les a produits que les tubercules de nouvelle formation, constituant autant d'individus indépendants, émettent de leur bord de nombreuses racines fines et indivises ; c'est ainsi du moins que les choses se passent généralement. Mais lorsque le tubercule nourricier n'a qu'un petit volume, on voit tantôt toutes les pousses qui en dérivent, tantôt quelques-unes d'entre elles seulement, se prolonger à leur base en une racine monstrueuse, gonflée de sucs, et dépassant en diamètre la base même de la pousse qui la produit. Il suffit de jeter les yeux sur les figures 15 et 16 pour voir l'énorme différence qui sépare cette racine des racines normales du vieux tubercule. J'ai constaté le même fait dans le Gladiolus communis. Cette racine atteint son complet développement au moment de la végétation la plus active de la plante mère, alors que le bourgeon, dont elle constitue une dépendance, commence à s'organiser en tubercule (fig. 16). Plus tard, lorsque le tubercule nourricier commence à s'épuiser, on la voit se couvrir vers sa base de larges rides qui l'envahissent peu à peu, et qui occasionnent la résorption active dont elle est le siège. Enfin le nouveau tubercule atteint sa maturité, et la racine qui l'a nourri de sa substance n'est plus qu'une dépouille aride qui se détache en laissant une faible cicatrice ; ses fonctions ne sont donc pas douteuses. Ne pouvant être assimilées en entier par les bourgeons encore trop jeunes, les substances que contient déjà ou qu'élabore la plante mère pendant la plus grande activité de sa végétation s'accumulent dans ce réservoir provisoire, y subissent peut-être une nouvelle préparation, et passent enfin, lorsque le vieux tubercule est flétri, dans le nouveau tubercule, but final de tout cet admirable appareil. Telles me paraissent être encore les fonctions qu'ont à remplir la racine napiforme et le tubercule basilaire de l'Ophrys apifera.
§ II. Développement du tubercule terminal.
Si je ne me fais illusion, l'exposé que je viens de donner de la germination et des premières phases de l'Ophrys apifera me paraît établir sur des bases certaines que le tubercule terminal de celle plante, et par suite d'une manière générale le tubercule des Ophrydées, n'est pas une sorte de cul-de-sac formé par une feuille, et où s'accumule la masse radiculaire d'un bourgeon, opinion à laquelle s'est arrêté M. Germain de Saint-Pierre (Bull. de la soc. bot. de France, n° de septembre 1855, p. 659), mais purement et simplement un entre-noeud renflé en tout ou en partie en tubercule. Il ne sera cependant pas inutile, tant pour confirmer l'opinion que j'adopte que pour jeter quelque lumière sur la structure complexe de cet organe et surtout de son pédicelle, d'en suivre pas à pas la formation sur les plantules issues de graines. Coupons suivant son axe une de ces plantes embryonnaires aussi avancée que possible, mais toutefois avant l'apparition du double mamelon dont j'ai parlé plus haut ; sa longueur est alors de 1 millimètre 1/2 à 2 millimètres. La gemme (fig. 10) se compose alors de deux feuilles rudimentaires, dont l'externe est le cotylédon. Au fond de cette double gaine se montre le mamelon, le point vital qui doit fournir les feuilles suivantes. Rien à cette époque ne trahit la marche anormale que doit suivre l'évolution de l'axe ; tout se montre parfaitement régulier et conforme aux lois ordinaires. Les feuilles ont un lien d'insertion manifeste, que nul ne s'aviserait de transporter ailleurs ; en un mot, le lieu de l'insertion apparente est bien dans la figure 10 celui de l'insertion réelle. Mais attendons un peu ; voici qu'au-dessous de ces feuilles et un peu par côté se dessine un trait presque insensible, circonscrivant partiellement un noyau, ne différant en rien, du reste, de la masse tigellaire. Ce trait de démarcation n'aboutit pas de part et d'autre à l'intervalle qui sépare le cotylédon de la feuille intérieure, il laisse, au contraire, de chaque côté une espèce d'isthme, qui relie le noyau au reste de la plantule. L'isthme interne, ou du côté concave, beaucoup plus large que l'autre, reçoit l'axe vasculaire de la tigelle. L'insertion des feuilles est-elle maintenant déplacée ? Évidemment non. L'insertion est la même que précédemment, et le trait délicat qui se dessine à peine ne longe pas la face interne du cotylédon, mais est tracé dans la substance même de la tigelle ; ou en d'autres termes et pour être plus clair, la partie placée à gauche de ce trait appartient à l'axe, à la tigelle, et non à la feuille externe. A plus forte raison, ne peut-on pas dire que la partie placée à droite du même trait, que le noyau est revêtu de ce côté par la base de la feuille interne. Encore une fois, précisons bien la question. Ce trait est-il la ligne de séparation de la base des deux feuilles, ou bien est-il imprimé dans la substance même de l'axe ? Cette dernière manière de voir me paraît avoir pour elle toute l'évidence désirable. Mais, avant de me faire cette concession, que mes adversaires y songent bien, là est le noeud du problème, derrière ce trait est le tubercule. Si on me l'accorde, il n'y a plus de cul-de-sac formé par une feuille, il n'y a plus de masse radicellaire constituant un renflement charnu. Si on me la refuse, je demanderai à mon tour par quelles permutations magiques l'insertion des feuilles, si évidente dans la figure précédente, se trouve maintenant transportée plus bas, presque au centre de la tigelle ; je demanderai enfin pourquoi cet isthme de gauche que j'ai invariablement retrouvé partout, et que rien n'expliquerait, si l'on ne veut y voir la base insertionnelle des deux feuilles. Jusqu'à ce que ces deux graves difficultés soient levées, et je ne vois rien qui puisse y amener, je regarderai le trait en question comme imprimé dans la masse même de la tigelle.
Mais ce trait devient un sillon ; ce sillon grandit, et se change en large lacune, dont les bords éraillés annoncent une destruction rapide des tissus voisins (fig. 12). En même temps, le noyau formé par la partie culminante de la tigelle s'isole davantage, prend la forme de plus en plus manifeste du tubercule futur, et, poussé par une force irrésistible, est enfin obliquement chassé hors de sa place primitive en entraînant avec lui, dans son expulsion, les diverses feuilles de la gemme. Sous sa pression, la paroi de l'espace lacunaire se distend, et forme au dehors le premier des deux mamelons déjà mentionnés, et dont le second est dû à une racine adventive (fig. 13). Enfin cette paroi crève pour laisser sortir la pointe jaunâtre du tubercule rudimentaire autour duquel elle constitue une gaine délicate.
Je relèverai ici une erreur que j'ai commise dans mon précédent travail sur les tubercules des Ophrydées. Faute d'avoir observé des bourgeons assez jeunes, je n'avais pas été témoin, comme dans le cas actuel, de la manière dont le tubercule se forme par l'isolement graduel de la partie culminante et centrale de l'axe, de la manière enfin dont s'opèrent et cette espèce de décortication du noyau central, et la formation de l'espace lacunaire qui le circonscrit en moyenne partie. Rien ne me faisant donc soupçonner son absence à un certain moment, j'ai pris cette lacune pour l'intervalle séparant l'axe du bourgeon et sa première feuille, ce qui m'a grossièrement fait méprendre sur l'insertion réelle de la base externe de cette feuille. Cette insertion se trouve, en effet, au même niveau que l'insertion de la seconde, là où se trouve cet isthme que la lacune n'a pas atteint, et que je prenais alors pour une soudure impossible à expliquer, et non de l'autre côté du tubercule à la naissance même de l'espace lacunaire, (Recherches sur les tubercules de l'Himantoglossum hircinum, Ann. des sc. nat., 4° série, t. III, fig. 1 et 2). D'après cela, dans la figure 4 du mémoire cité, la bourse contenant le noyau tuberculaire n'est pas formée par la base de la première feuille du bourgeon dilatée en cul-de-sac, mais bien par la couche externe de l'axe détachée du reste de la masse axile par le mécanisme que je viens d'exposer. Enfin, dans la figure 6, le point c est réellement l'insertion de la première feuille, et la gaine bd cd est, comme la bourse précédente, la dépouille superficielle du rameau tuberculaire. Avec ces modifications qui rapportent à sa véritable origine l'espèce de coléorhize qui revêt la base pédicellaire du tubercule, et rétablissent en son lieu vrai l'insertion de la première feuille du bourgeon, on voit que le tubercule est formé non par l'extrémité du second entre-noeud, comme !'exigeaient les idées mal fondées que je viens de rectifier, mais bien par l'extrémité du premier.
Mais cherchons à jeter quelque jour sur la formation si exceptionnelle du tubercule de l'Ophrys. Dans une gemme dont rien ne trahit encore au dehors les profondes perturbations internes, la partie culminante et centrale s'isole, disons-nous, du reste de la tige, se conglobe en noyau, et déserte sa place originelle, tandis que les tissus dont elle faisait d'abord elle-même partie se fondent, s'évanouissent pour ainsi dire à son approche, et lui abandonnent un libre passage. Les causes de ces phénomènes complexes m'échappent, et je ne chercherai pas à expliquer ce qui peut-être est inexplicable. Je m'arrêterai simplement à démontrer, par un exemple pris sur une autre plante, que, lorsqu'il s'agit de conserver l'étincelle de vie confiée à un germe, la nature a parfois des procédés bien autrement éloignés de ses voies ordinaires. C'est dans la germination du Colchique (Colchicum autumnale) que je prendrai mon exemple. Semées en mai, les graines ont germé le mois de février suivant, et ont produit en mars des plantules composées d'un cotylédon encore engagé dans la graine, d'une feuille subulée et d'une radicule indivise (fig. 17). La démarcation de la radicule et de l'axe ascendant se dessine par une diminution assez faible, mais subite, dans le diamètre respectif des deux organes. Au niveau de cette démarcation ou, en d'autres termes, au niveau du collet, la base engrainante de la feuille subulée enveloppe un mamelon très menu. Jusqu'ici tout est donc parfaitement normal. En avril, aucun nouvel organe n'a apparu à l'extérieur. Arrachons alors avec précaution cette plante délicate ; la radicule en un point de sa longueur est oscillante, comme si un coup maladroit de boulette l'avait endommagée. En outre, sa moitié supérieure est flasque et d'un aspect louche particulier. En l'observant par transparence, on croirait voir un corps étranger et blanc logé sous son épiderme diaphane ; et c'est en effet ce qui a lieu. En fendant cet épiderme, on trouve (fig. 18), au lieu du tissu de la radicule, un cylindre blanc occupant environ la moitié supérieure de la radicule, et se continuant plus haut avec la feuille subulée. Inférieurement, ce cylindre se termine un peu en pointe, et se rattache à un cordon capillaire, dirigé d'abord de bas en haut, puis rétrogradant de haut en bas pour plonger enfin dans la partie intacte de la radicule dont il constitue l'axe vasculaire. Le cylindre lui-même se compose (fig. 17) de la base de la feuille subulée, dont l'étroit canal s'élargit un peu inférieurement pour recevoir un second cylindre beaucoup plus court et épais, constituant le premier réservoir de fécule, le premier tubercule. Au sommet de ce tubercule cylindrique se montre un faible bourgeon. Nous avons donc encore ici un exemple d'un tubercule s'organisant aux dépens de l'extrémité de l'axe. Mais quelle route singulière cette gemme terminale ne suit-elle pas pour s'enfouir plus profondément ! Sa base était d'abord au niveau du collet de la plante ; elle est maintenant à plusieurs centimètres plus bas, au milieu de la radicule ! Il a donc fallu à cette gemme se débarrasser d'abord de ses connexions avec la feuille cotylédonnaire abandonnée en arrière, puis, par un mouvement rétrograde, plonger profondément dans la radicule dont le tissu cellulaire disparaît à son approche, mais dont l'épiderme persiste ainsi que le faisceau vasculaire, pour lui former, le premier, un fourreau protecteur, le second, un cordon ombilical, qui la met en rapport avec la partie intacte de la radicule. C'est ce cordon que la gemmule entraîne dans sa descente en le doublant en boucle, et qui, développé, atteindrait précisément le point de départ, le collet. La gemme conserve encore quelques vestiges de son évulsion violente de sa place primitive ; sa base est rugueuse, éraillée, et on y trouve appendu un filament vasculaire, qui ne peut être qu'un débris de la nervure du cotylédon (fig. 19). Je ferai observer que l'invagination de la gemme dans la radicule n'est pas un fait accidentel dû à la situation de cette dernière sur la même verticale que la gemme doit parcourir; car, quelque flexueuse que soit la radicule, la gemme en suit fidèlement les contours. Je ferai remarquer encore que, pour accompagner ainsi le bourgeon terminal dans son émigration, la base de la feuille subit un allongement considérable, à l'époque même où sa partie supérieure et aérienne reste complètement stationnaire. Tôt ou tard le fourreau, formé par la radicule envahie, devient cependant trop étroit pour contenir le corps étranger ; il se fend alors irrégulièrement, et abandonné à elle-même la gemme qui, suffisamment enfouie, peut désormais mûrir son tubercule en sécurité. Les tubercules adultes du Colchique présentent une tendance analogue à s'enfouir plus profondément. On sait que ces tubercules émettent fréquemment à leur base un prolongement, parfois assez long, destiné à maintenir à une profondeur suffisante le bourgeon inférieur, le plus important des deux que porte le tubercule. De ce processus gemmifère au noyau terminal de l'Ophrys suspendu à un long pédicelle, il n'y a pas loin ce me semble. Le but est toujours le même, la conservation d'une gemme. Mais pour atteindre ce but, les moyens diffèrent dans leurs détails ; car « chaque espèce à tige souterraine, chaque espèce bulbeuse, a pour ainsi dire une manière de végéter, des habitudes qui lui sont propres, je dirais presque des moeurs (Aug. de Saint-Hilaire, Morphol. végét., p. 125). »
Après cette digression dans le domaine de la botanique comparée ; après avoir constaté, par un exemple où tout est évident, l'évulsion d'une gemme terminale de sa place originelle, sa pénétration dans les profondeurs de la radicule, et la résorption des tissus de cette dernière pour lui livrer passage, la formation du tubercule terminal de l'Ophrys apifera n'a plus rien qui puisse nous étonner. Suivons donc dans son trajet le noyau expulsé du centre de la plante. La couche externe de la tigelle vient de crever sous sa pression, et forme une courte gaine à sa base. L'isthme interne, plus accentué, dessine déjà assez bien un rudiment de pédicelle (fig. 5). La feuille cotylédonaire est abandonnée en arrière ; mais la feuille suivante continue à accompagner la gemme, comme elle le fait dans le Colchique. La chose est évidente pour la face externe de celle feuille, et, malgré une soudure intime entre l'axe ou le pédicelle et la base interne de la même feuille on ne peut s'empêcher d'admettre la décurrence de cette base sur l'axe, et de transporter son insertion au niveau de la base externe. L'exemple du Colchique et une étude scrupuleuse à la loupe ne me laissent pas le moindre doute à ce sujet. Plus tard, la gemme produit une seconde feuille qui concourt à son tour à la formation du pédicelle ; et si l'on attend que la plantule ait acquis à peu près tout son développement (fig. 6), on trouve, pour le tubercule et son pédicelle, la structure représentée dans tous ses détails dans la figure 14, structure fort aisée à expliquer avec le secours des notions qui précèdent. Dans celle figure, les lignes pleines sont le résultat de l'observation, les lignes ponctuées sont théoriques. La feuille cotylédonaire a, abandonnée en arrière, est insérée en b et en c. L'intervalle c, qui ressemble à une soudure, est l'isthme externe dont j'ai parlé plus haut, ou l'intervalle épargné par le sillon lacunaire qui s'est formé au moment de la délimitation du tubercule. La gaine of, ch, n'appartient pas au cotylédon, mais comme je l'ai dit, à la tigelle t. Elle est le résultat de la rupture de la couche placée à gauche de la lacune dans la figure 12. La feuille suivante i est adhérente en c avec la base du colylédon. Les figures 11, 12, 13 et 5, montrent l'origine de cette adhérence qui n'a rien d'exceptionnel, puisque la région où elle se trouve n'est autre chose que l'ensemble des bases primitives des deux feuilles. Pour concevoir le prolongement de la seconde feuille au-dessous de cette région, il faut se figurer, dans la base insertionnelle primitive de cette feuille, un développement pareil à celui dont le Colchique nous offre un si bel exemple ; il faut se figurer, dis-je, qu'après la production de sa partie aérienne, la feuille recommence, mais en sens inverse, son élongation ; ce qui produit la partie comprise entre c et k. La troisième feuille l est libre jusqu'en m ; plus bas, de m en k, elle se solide avec la seconde. On peut encore se rendre compte de cette soudure en admettant que, tant que la gemme n'a pas atteint le niveau m, l'élongation rapide de la base de cette feuille n'a pas lieu ; mais qu'à partir de ce moment, cette élongation commence, et se fait en commun avec celle de la feuille précédente. Parfois, mais plus rarement, la soudure mk n'existe pas, et les deux feuilles sont parfaitement libres jusqu'au point k, lieu de leur insertion, ce qui prouve suffisamment que, dans le cas actuel, il faut regarder la couche mk comme la somme des bases des deux feuilles ; voilà pour la paroi externe du canal du pédicelle.
La paroi interne n'est pas plus difficile à expliquer ; la feuille la plus intérieure conserve des traces manifestes de sa décurrence dans toute l'étendue du canal, et l'on rapporte, sans indécision, son insertion au point n, ce qui d'ailleurs ne saurait être autrement. La feuille précédente, plus intimement soudée, ne laisse pas de trace sensible de la décurrence de sa base. Cependant comme, d'une part, on trouve parfois, ainsi que je l'ai dit, cette feuille librement insérée en k ; comme d'autre part la première évolution du pédicelle laisse assez bien constater cette décurrence, j'admettrai que cette feuille vient se relier, comme l'autre en n, au sommet des tubercules; et je regarderai comme l'expression de la vérité les deux lignes théoriques qui partant de s aboutissent en n. Quant au reste du pédicelle, il est évidemment formé par l'axe lui-même, par le second entre-noeud de la plantule, le premier étant le tubercule tigellaire t. Le même faisceau vasculaire les parcourt sans interruption l'un et l'autre, et s'évanouit dans le voisinage de la gemme du tubercule. Si je suis parvenu à me faire suffisamment comprendre dans cette aride démonstration, on admettra, je crois, sans difficulté que l'organe complexe que je viens de décrire se compose tout simplement du second entre-noeud de la plantule renflé à l'extrémité et muni d'une gemme campylotrope dont les premières feuilles se soudent, par une de leurs faces, entre elles et avec l'axe qui les porte. Il est inutile d'ajouter que les tubercules provenant de bourgeons axillaires reconnaissent absolument la même structure et le même mode de formation. Ils sont, par rapport à des axes secondaires, ce que le tubercule terminal est par rapport à l'axe primaire.
§ III. Tubercules palmés.
Les tubercules palmés des Ophrydées fournissent le principal argument en faveur de l'opinion contraire à celle que je soutiens. Aux yeux de mes adversaires, les prolongements de ces tubercules sont de véritables racines, qui attestent la nature radiculaire de la masse qui les produit. Il importait donc d'accorder une attention toute spéciale à ces tubercules : c'est ce que j'ai fait en prenant pour sujet de mes recherches l'Orchis divaricata. J'avouerai tout d'abord que ces prolongements n'ont rien d'essentiel, à la structure anatomique près, qui m'empêche de les regarder comme des racines ; je confesserai même qu'ils en remplissent les fonctions, qu'ils absorbent parfaitement les liquides ambiants. Dans une faible dissolution de cyanoferrure de potassium, j'ai fait plonger tantôt simplement les racines adventives de l'Orchis, tantôt au contraire les prolongements du tubercule. Puis fendant la plante dans toute sa longueur, et promenant sur la section un pinceau imbibé d'une dissolution d'un sel de fer, j'ai obtenu, dans les deux cas, une coloration due au bleu de Prusse également intense et également propagée, lorsque la durée de l'immersion et l'étendue des surfaces étaient à peu près les mêmes. Je dirai plus, les tubercules ovoïdes et indivis absorbent les dissolutions salines avec la même facilité, pourvu qu'ils soient jeunes, ou, pour mieux dire, pourvu qu'ils appartiennent à la formation de l'année. Mais la fonction ne fait pas l'organe, et il le faut bien puisque nous avons vu la plantule embryonnaire de l'Ophrys apifera arriver, quoique sans radicule, à un volume énorme relativement à son point de départ. Elle absorbait donc, elle absorbait par sa tigelle, par son premier tubercule.
Une preuve, à mon avis sans réplique, en faveur de la nature caulinaire du tubercule terminal de l'Ophrys, c'est son mode de formation au sommet de la plantule. Franchement, qui s'aventurerait à regarder comme un faisceau de racines agglutinées la partie culminante de l'axe qui graduellement se délimite, se façonne en noyau, et s'échappe en entraînant la gemmule ? Eh bien, ce mode de formation se retrouve, sans le moindre changement, dans l'Orchis divaricata. Je n'ai pas été témoin, il est vrai, de la germination de cette plantule ; mais heureusement il n'est pas nécessaire d'assister aux premiers débuts des Ophrydées pour trouver des tubercules terminaux, puisqu'il s'en forme plusieurs années de suite. J'ai déjà signalé la métamorphose de la sommité de l'axe en tubercule dans les jeunes pousses d'Himantoglossum hircinum, et pour rédiger ces pages, je l'ai observée encore dans les Ophrydées suivantes : Ophrys apifera, Ophrys myoides, Orchis divaricata, Orchis laxiflora, Orchis fusca, c'est-à-dire dans toutes les Ophrydées que j'ai pu observer assez jeunes. Ce fait paraît donc général, et si quelque chose m'étonne, c'est que les auteurs qui se sont occupés de la germination de ces plantes ne l'aient pas déjà fait connaître. J'ai trouvé même des pousses d'Orchis laxiflora ayant à leur base la grosseur du doigt, c'est-à-dire ayant atteint leur plus grand développement, dont le sommet de l'axe, au lieu d'une grappe de fleurs, avait produit un tubercule. Dans ces pousses, le bourgeon supérieur ou le troisième, au lieu de produire immédiatement un tubercule, se développait en une tige aussi vigoureuse et aussi longue que la tige mère, et, après avoir fourni un nombre assez considérable de feuilles, suspendait tout à coup son évolution pour donner naissance à un tubercule terminal ; ce qui prouverait, s'il en était besoin, que les bourgeons axillaires se comportent comme la gemme terminale.
La figure 20 reproduit de grandeur naturelle une plantule d'Orchis divaricata, munie d'un tubercule terminal en voie de formation. On voit sous la troisième écaille un mamelon jaunâtre dû à l'apparition de ce tubercule. Le tubercule qui a produit cette pousse est indivis, napiforme, comme l'est celui du Platanthera. Dans les plantules adultes, il est, au contraire, bifurqué à une distance plus ou moins grande du bourgeon. Quelquefois même l'une des bifurcations ou toutes les deux se dédoublent, ce qui produit 3-4 prolongements radiciformes. La figure 21 est une section de la partie supérieure de la jeune plante. On voit que le mécanisme par lequel se produit le tubercule est le même que dans l'Ophrys. C'est toujours une portion du sommet de l'axe qui s'isole, et s'ouvre un passage à travers les tissus qui l'emprisonnent. Le noyau tuberculaire est alors globuleux, sans aucune apparence de prolongement radiciforme, et l'on ne peut s'empêcher de proclamer son identité avec ceux de l'Ophrys apifera. Hors de la pousse, ce globule continue à grossir tout en conservant sa forme ovoïde ; puis à sa base, on voit poindre un mamelon jaunâtre, qui en s'allongeant finit par donner au tubercule l'aspect napiforme. Les bourgeons axillaires suivent dans leur évolution la même marche. L'axe du bourgeon, par une sorte de décortication, produit dans son intérieur un noyau ovoïde, ainsi que je l'ai déjà dit à propos de l'Ophrys apifera. Ce noyau grossit, perce la couche externe qui l'enveloppe et s'engage dans cette ouverture comme dans une gaine. Son volume est déjà assez considérable, lorsque se montrent deux mamelons qui s'allongent et prennent l'aspect de deux racines. En résumé, et pour éviter des redites fastidieuses, les bourgeons axillaires comme les gemmes terminales suivent, pour s'organiser en tubercules, absolument la même marche dans l'Ophrys apifera et dans l'Orchis divaricata, et avoir démontré d'après ce mode de formation la nature des tubercules de la première plante, c'est avoir démontré la nature de ceux de l'autre ; c'est avoir établi que les tubercules palmés, du moins dans leur partie indivise, sont formés par l'extrémité hypertrophiée d'une tige on d'un rameau. Quant aux prolongements que ces tubercules émettent, voici ce qui me paraît le plus probable. Ils ont, ai-je dit, l'aspect des racines ; mais cette ressemblance ne se maintient pas pour la strucure interne. L'axe d'une racine est occupé par une rangée circulaire de petits faisceaux vasculaires ; dans les prolongements radiciformes des tubercules, ces mêmes faisceaux sont distribués çà et là sans aucun ordre, comme ils le sont dans le corps du tubercule, comme ils le sont enfin dans une tige de Monocotylée. En outre, l'ensemble des faisceaux qu'on observe dans le corps du tubercule se retrouve dans les prolongements. J'ai compté 26 faisceaux dans la masse indivise d'un tubercule par deux fois bifurqué, et j'ai retrouvé ce nombre 26 pour somme des faisceaux observés à l'extrémité des quatre divisions. Je ferai observer enfin que l'amas féculent ne dépasse pas le corps du tubercule, que le tissu cellulaire des prolongements ne renferme pas de grains amylacés. Je m'explique cette structure comme il suit. Un tubercule se compose d'un double système d'organes élémentaires, un tissu cellulaire féculent, et un tissu vasculaire plongeant par colonnes dans le premier pour lui former comme une charpente, si je peux me servir de cette expression. Si l'élongation des colonnes vasculaires ne marche pas plus rapidement que le dépôt féculent qui s'effectue dans leurs intervalles, le tubercule reste ovoïde; si, au contraire, cette élongation est plus rapide, que la formation amylacée, si elle conserve une même activité quand celle formation se ralentit, il arrive un moment où les colonnes vasculaires ne peuvent plus se relier entre elles en une seule masse, et alors elles continuent leur trajet seules, par faisceaux plus ou moins nombreux, suivant leur divergence primitive. De là la masse indivise et féculente et ses prolongements sans fécule, simples partitions appauvries de cette masse. Si cela est, un sol plus substantiel, en rendant plus abondante la production de fécule, doit tendre à effacer plus ou moins ces prolongements. C'est une expérience à faire.
Si, malgré la différence de structure que j'ai signalée, on veut voir de vraies racines dans ces prolongements, voici l'idée que je proposerais : On a vu que letubercule basilaire de l'Ophrys produit une racine napiforme considérable venant en aide au tubercule pour élaborer la gemme terminale ; j'ai dit que cette racine était parfois tellement continue avec le tubercule, qu'on prendrait cet ensemble pour un organe unique. Que l'on compare le tubercule basilaire muni d'une racine napiforme de la figure 7, avec le tubercule d'Orchis divaricata de la figure 20 : quelle, différence trouve-t-on ? Aucune. Dans la figure 16 relative au Safran, supposons au tubercule actuel le volume qu'il doit acquérir plus tard ; dépouillons-le de ses feuilles pour ne lui laisser que la gemme supérieure ; faisons disparaître l'étranglement qui le sépare de son énorme racine napiforme, et nous aurons le tubercule du Platanthera bifolia. Au tubercule globuleux de l'Ophrys apifera, attachons deux racines pareilles, nous aurons le tubercule de l'Orchis divaricata. Dans cette hypothèse, le tubercule palmé serait le tubercule globuleux, plus des racines adventives analogues à celle que j'ai fait connaître dans le Safran. Il n'y aurait d'ailleurs rien d'étonnant à avoir ici des tubercules sans racines, là au contraire avec des racines ; car n'oublions pas que le tubercule tigellaire de l'Orchis morio est absolument sans racines ; que celui de l'Ophrys apifera en a une considérable ; que, dans cette dernière espèce même, si le tubercule tigellaire en a une, le tubercule basilaire de seconde année en a deux, et que le tubercule terminal n'en a point. Malheureusement, je le répète, l'anatomie de ces prolongements n'est pas en rapport avec cette interprétation.
Conclusions.
1° La tigelle de l'embryon macropode de l'Ophrys apifera forme le premier tubercule de celle plante.
2° La jeune plante est dépourvue de radicule, et se compose à son début du tubercule tigellaire surmonté d'une gemme, rappelant ainsi, de la manière la plus exacte, les tubercules qui doivent s'organiser plus tard aux dépens, soit de la gemme terminale, soit des bourgeons axillaires.
3" Le tubercule tigellaire produit tôt ou tard une racine adventive qui n'a rien de particulier.
4° La gemme terminale s'organise en tubercule, c'est-à-dire que la portion terminale de l'axe se conglobe en noyau, et s'ouvre une issue violente au dehors en entraînant la gemme.
5° Ce noyau tuberculaire est suspendu à l'extrémité d'un long pédicelle formé d'un côté par l'axe de la plante, et de l'autre par les premières feuilles de la gemme soudées avec cet axe. Le but du pédicelle est d'amener la gemme tubéreuse à une profondeur suffisante pour y passer l'hiver en sécurité.
6° Le tubercule ainsi formé, et que j'appelle terminal, a la même structure anatomique que le tubercule tigellaire.
7° Il ne reproduit pas la plante, il la continue.
8° La pousse issue l'année suivante de ce tubercule produit à sa base un renflement tubéreux, rappelant à s'y méprendre le tubercule tigellaire. J'appelle ce renflement tubercule basilaire. La sommité de la pousse produit en même temps un tubercule terminal en tout pareil au premier.
9° Du tubercule basilaire s'échappent deux racines adventives, dont l'une placée à sa base est napiforme, et offre les plus grands rapports de structure et de fonctions avec une racine pareille que j'ai fait connaître dans le Safran.
10° Après un nombre encore indéterminé d'évolutions annuelles pareilles, l'axe issu de la graine se termine en tige ordinaire, stérile et sans tubercule terminal. Les bourgeons axillaires sont alors chargés de reproduire la plante, de la multiplier, et d'amener enfin la forme florale après plusieurs générations par gemmes.
11° Si l'on suppose que cet axe primitif produise en une saison tous les tubercules qu'il produit à sa base et à son sommet par périodes annuelles, on aura la structure d'un chaume d'avena bulbosa, dont les entre-noeuds inférieurs forment un chapelet de tubercules.
12° Ce mode d'évolution paraît être général dans les Ophrydées ; je l'ai constaté en tout ou en partie chez toutes les Ophrydées que j'ai pu observer assez jeunes.
13° Les tubercules ovoïdes des Ophrydées, soit axillaires, soit terminaux, résultent également d'un noyau évulsé de la partie centrale et terminale d'un axe. La couche qui revêt ce noyau est la partie de l'axe éliminée de cette formation ; c'est elle qui, sous la pression du noyau, se rompt, et forme la gaine qu'on trouve à la base du pédicelle. Par erreur, j'ai rapporté d'abord cette gaine à la première feuille de la gemme.
14° Ce noyau, ce tubercule ne peut, à cause de son origine, avoir rien de commun avec des racines ; il ne peut non plus, à cause du mécanisme de sa formation, être contenu dans un sac, un éperon, formé par les bases dilatées de ses premières feuilles.
15° Le pédicelle qui le supporte est formé d'un côté par l'axe même dont le tubercule est l'extrémité, et du côté opposé par les premières feuilles de la gemme campylotrope soudées avec l'axe par une de leurs faces, ainsi que je l'ai exposé dans mon précédent travail, dont toutes les conclusions sont maintenues après la rectification mentionnée au n° 13.
16° Les tubercules palmés, ayant la même origine et le même mécanisme de formation que les tubercules ovoïdes, reconnaissent la même nature que ces derniers.
17° Leurs prolongements radiciformes sont ou de simples partitions résultant de l'élongation des colonnes vasculaires plus rapide que la formation du tissu féculent ; ou bien encore des racines adventives analogues à la racine napiforme observée dans l'Ophrys apifera et dans le Safran.
18° Leur structure anatomique n'étant pas celle des racines, la première opinion me paraît cependant plus probable.
source : Annales des Sciences Naturelles, 4ème série, Tome V, Cahiern° 3.