L'ANATOMIE DES ORGANES REPRODUCTEURS ET SUR LE DÉVELOPPEMENT DES MYRIAPODES
DEUXIÈME PARTIE
CHILOPODES.
CHAPITRE PREMIER.
ORGANES DE LA GÉNÉRATION.
1. Organes femelles.
Ovaire. — Mes recherches embrassent les genres Lithobius, Scutigera, Scolopendra, Cryptops et Geophilus. Dans tous, l'organe préparateur et éducateur des ovules forme un long sac impair, s'étendant de l'extrémité postérieure du corps jusque vers l'origine des glandes salivaires et vénénifiques. Cet organe est placé à la partie dorsale, au-dessus du tube digestif; tandis que, chez les Chilognathes, il est situé au-dessous de ce tube. Le sac ovarique est plongé dans du tissu adipeux de forme variable, et se trouve maintenu en place par de nombreux ramuscules trachéens qui se répandent dans l'épaisseur de sa paroi inférieure. En l'ouvrant longitudinalement, on reconnaît que les ovules ne se développent que sur cette paroi occupée par un stroma unique ou placentaire, qui s'étend presque d'un bout à l'autre du sac (fig. 9). Comme chez les Chilognathes, les ovules sont enfermés chacun dans une capsule ovarique, rattachée au placentaire par un très court pédicule. Ils sont fort inégalement développés d'un bout à l'autre du stroma ovuligène, les plus gros pêle-mêle avec les moins avancés. A mesure que leurs capsules se rompent, les ovules mûrs et libres s'entassent dans le sac ovarique qu'ils boursouflent irrégulièrement. Pas plus que chez les Chilognathes, ce sac ne pouvant être comparé aux tubes ovariques des Insectes, je continuerai à le désigner par le nom de sac ovarique, en réservant le nom d'ovaire au stroma ovuligène unique qui double sa paroi inférieure. L'ovaire est donc unique chez les Chilopodes, contrairement à ce qui a lieu chez les Chilognathes. M. Léon Dufour, qui a donné l'anatomie du Lithobius forcipatus (1), est porté à croire que l'ovaire (sac ovarique) de cette espèce est divisé intérieurement en deux loges par un diaphragme longitudinal. Il n'en est rien, pas plus chez les Lithobies que chez les autres Chilopodes. D'ailleurs, l'existence d'un seul stroma ovuligène s'oppose évidemment à la présence d'un diaphragme. A cause de l'uniformité de cet appareil chez les divers Chilopodes, il est inutile d'en donner une description particulière pour chaque genre.

Fig 9
Portion ouverte du sac ovarique du Scolopendra complanata pour montrer l'ovaire unique.
Oviductes.—Le stroma ovuligène s'arrête à une certaine distance de l'extrémité postérieure du corps; mais le sac ovarique se prolonge encore un peu en se rétrécissant, et se termine, enfin, soit par un oviducte double (fig. 10), dont les deux branches forment une anse qui embrasse le rectum (Lithobius, Scutigera) ; soit par un oviducte simple (Scolopendra, Cryptops, Geophilus), qui, d'abord supérieur au rectum, plonge enfin au-dessous de lui, de sorte que, dans le cas d'un oviducte simple comme dans celui d'un oviducte double, l'orifice génital est inférieur à l'anus (fig. 11, 12 et 13).

Fig 10
Organe génitaux femelles du Scutigera araneoides.
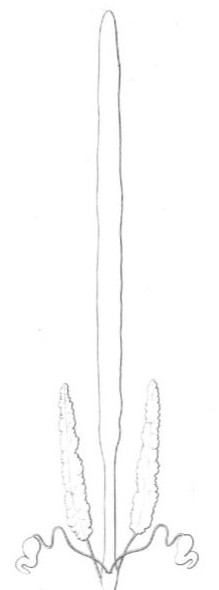
Fig 11
Organes génitaux femelles du Scolopendra complanata.

Fig 12
Organes génitaux femelles du Cryptops Savignyi.

Fig 13
Organes génitaux femelles du Geophilus Gabrielis.
M. Léon Dufour n'a pas vérifié chez les Lithobies la manière dont l'ovaire se comporte en s'approchant de la vulve ; il admet, d'après les observations et les figures de M. Treviranus, que cet ovaire se termine par un oviducte simple et dilaté à son extrémité postérieure. M. Straus-Dürckheim est plus heureux lorsqu'il décrit cet ovaire comme produisant deux oviductes qui reçoivent le rectum entre eux (2). Quant au Scutigera araneoides, M. Léon Dufour, qui s'est également occupé de l'anatomie de cette espèce (3), ne parle pas de ses oviductes. Comme chez les Lithobies, il y a deux oviductes qui embrassent le rectum dans leur anse.
Réceptacles séminaux.—Chez les Lithobies, ces réceptacles ont été pris comme des réservoirs des quatre glandes génitales postérieures par M. Léon Dufour, qui admet que les déférents de ces glandes s'insèrent au bout antérieur des réservoirs. Chez les Scutigères, ce savant anatomiste n'a pas aperçu les réservoirs spermatiques. Le disque lenticulaire opaloïde, qu'il décrit comme glandes sébacées, est, en effet, formé par les glandes accessoires et non, comme le pense M. Siebold (4), par les réceptacles séminaux eux-mêmes. M. Stein a parfaitement reconnu ces réservoirs chez le Lithobius forcipatus et le Geophilus subterraneus.
J'ai trouvé ces deux réceptacles séminaux cher tous les Chilopodes. Ils sont constamment placés côte à côte sur la ligne médiane, dans les derniers anneaux du corps, au-dessous du tube digestif, et se composent de deux utricules de forme variable, d'un blanc amylacé et hyalins sur les bords. En les déchirant, on reconnaît qu'ils sont formés d'une tunique transparente fort délicate, et que leur contenu consiste en un noyau blanc composé de myriades de Spermatozoïdes capillaires, très longs et enchevêtrés en feutre serré. Chaque utricule se termine par un déférent capillaire, un peu rigide et tortueux. Ces réservoirs sont cylindriques, arrondis aux deux bouts, chez le Lithobius orcipatus; piriformes, chez le Scutigera araneoides (fig. 10) ; en massue, dont la partie étroite est capricieusement contournée, chez le Scolopendra complanata (fig. 11) ; en massue droite, chez les Cryptops hortensis et Savignyi (fig. 12); ovulaires, chez les Geophilus electricus, Ilicis convolvens (5) ; sphériques, chez le Geophilus Gabrielis (fig. 16).

Fig 16
Organes génitaux mâle du Scolopendra complanata.
La paroi supérieure du vestibule génital porte, à une très petite distance de la vulve, deux petits mamelons charnus, placés sur la ligne médiane l'un à côté de l'autre. C'est dans ces mamelons que débouchent les déférents des réceptacles séminaux.
Dans toutes mes observations, et j'en ai fait à toutes les époques de l'année, j'ai constamment trouvé les réservoirs séminaux des Chilopodes adultes remplis par un noyau de spermatozoïdes. C'est apparemment une pareille observation qui a porté M. Stein à annoncer cette circonstance fort extraordinaire, qu'on trouve dans les réceptacles séminaux des Lithobies et des Géophiles, sans accouplement préalable, des spermatozoïdes qui se développent dans ces organes en suivant les mêmes phases d'évolution que dans les testicules des mâles. Le fait annoncé était par trop exceptionnel pour ne pas me faire désirer de m'en convaincre par mes propres recherches. A cet effet, j'ai étudié la formation des spermatozoïdes dans les testicules des mâles ; j'ai décrit, dessiné les vésicules épithéliales dans lesquelles se développent les filaments spermatiques, et, après m'être familiarisé avec les diverses formes de ces vésicules, j'ai soumis les réservoirs des femelles à un examen comparatif. Sans entrer dans des détails fastidieux, je me bornerai à dire que, dans ces réservoirs, j'ai toujours trouvé des spermatozoïdes complètement développés, et jamais rien qui rappelât de près ou de loin les vésicules mères des testicules. D'ailleurs les spermatozoïdes des réceptacles étaient, la plupart du temps, rigides et immobiles, preuve manifeste qu'ils provenaient d'un résidu non employé à la fécondation précédente, et non d'une procréation sur place. Pas un seul Chilopode, quelle que fût l'époque de mon examen, n'a fait exception à cette règle. Je me crois donc fondé à conclure que l'assertion de M. Stein est erronée.
Glandes accessoires.—A sa terminaison, le canal vecteur des ovules reçoit les déférents de deux ou de quatre glandes sur la nature desquelles les auteurs sont loin d'être d'accord. Pour M. Treviranus, les grappes sécrétoires qui accompagnent les oviductes des Lithobies ne sont que des masses adipeuses ; pour M. L. Dufour, elles sont destinées à revêtir les oeufs d'une sorte de vernis ; en d'autres termes, ce sont des glandes sébacées. M. Straus-Durckheim, considérant que ces organes se trouvent également chez les deux sexes, admet que les glandes qui terminent l'appareil génital mâle ou femelle des Lithobies et des Scolopendres ne sont autre chose que des organes urinaires. J'ai soumis les glandes de la Scolopendre à l'action de l'acide azotique et de l'ammoniaque sans jamais obtenir la moindre trace de muréxide. En traitant au contraire de la même manière les vaisseaux de Malpighi, et surtout le produit blanc qu'ils déversent dans le rectum, j'ai toujours obtenu une belle couleur rose. L'acide urique est donc sécrété par les vaisseaux de Malpighi, et les glandes terminales de l'appareil génital ont un autre usage que celui que leur attribue M. Straus.
Leurs déférents n'ont aucun rapport avec les conduits séminaux des réservoirs. On ne peut donc pas les considérer comme sécrétant un liquide destiné à préserver les spermatozoïdes de la dessiccation et à les maintenir en bon état. D'ailleurs elles rempliraient fort mal leurs fonctions ; car, comme je l'ai déjà dit, les spermatozoïdes observés dans les réceptacles sont le plus souvent complètement rigides et morts. Elles me paraissent destinées à revêtir les oeufs d'un léger enduit, à les agglutiner, et surtout à faciliter l'accès du sperme dans les organes de la femelle, en faisant éclater les spermatophores que nous verrons déposer par les mâles.
Chez les Lithobies et les Scutigères, on trouve deux paires de ces glandes ; chez les autres Chilopodes, il n'y en a qu'une seule paire. Elles sont toujours placée au-dessous du tube digestif. Dans le cas de quatre glandes, les déférents des deux du même côté se rendent ensemble à un orifice commun, qui déverse leur contenu dans le vestibule génital à une très petite distance de son orifice. S'il n'y en a que deux, leurs déférents débouchent à part, l'un à droite, l'autre à gauche de !a terminaison de l'oviducte.
Lithobius forcipatus.
Les glandes accessoires sont allongées, lancéolées, tantôt diaphanes, tantôt d'un blanc amylacé, suivant leur degré de turgescence. Leur contour est vésiculeux et comme festonné. Les deux paires sont inégales : l'interne est beaucoup plus petite.
Scutigera araneoides (fig. 10).
M. L. Dufour a vu chez cette espèce un disque lenticulaire semi-diaphane ou opaloïde, se terminant par un gros pédicule ; mais la seconde paire a échappé au savant anatomiste, ainsi que les réceptacles terminaux.
Les glandes internes sont diaphanes, minces, discoïdes, et fournissent chacune plusieurs conduits capillaires groupés en faisceau, et se réunissant bientôt en un déférent commun. Les deux disques sont placés côte à côte, et adhèrent par leur côté interne aux réservoirs spermatiques placés sur la ligne médiane. C'est probablement cette adhérence qui a mis en défaut l'habileté ordinaire de M. L. Dufour. Ces mêmes disques adhèrent par leurs bords externes aux glandes de seconde paire. Celles-ci sont allongées, lancéolées, d'un blanc amylacé, et festonnées sur les bords : tel est du moins leur aspect au mois d'août. A d'autres époques, elles sont hyalines, et parfois si appauvries, qu'elles peuvent facilement passer inaperçues.
Chez les autres Chilopodes, on ne trouve plus qu'une seule paire de glandes.
Scolopendra complanata (fig. 11).
Elles sont étroites, allongées et vésiculeuses sur les bords, comme chez les Lithobies.
Cryptops Savignyi et C. hortensis (fig. 12).
Les glandes accessoires prennent ici une forme tout à fait insolite. Elles se composent de deux vaisseaux d'une excessive ténuité et d'une longueur démesurée, car ils s'étendent depuis les derniers segments jusque vers la base des glandes salivaires. Chacun porte sur sa face interne, et à des distances égales, une quinzaine de petites vésicules rondes, blanches et sessiles. Ce double cordon de vésicules est enchâssé dans le tissu adipeux au-dessous du tube digestif. Bien que chaque vaisseau, à cause de sa ténuité, soit difficile à démêler au milieu du tissu adipeux avec lequel il se confond par sa coloration, on peut cependant l'isoler entièrement en suivant de proche en proche les vésicules qu'il porte, et qui, malgré leur exiguïté, se dessinent parfaitement bien par suite de leur teinte d'un beau blanc. Dans leur partie terminale, les deux vaisseaux deviennent blancs, parce qu'ils sont remplis d'une humeur d'un blanc mat où flotte une pulviscule à grains transparents d'une extrême ténuité. Les vésicules renferment la même humeur.
Geophilus Gabrielis (fig. 13).
Cette espèce a deux glandules étroites, allongées, à parois vésiculeuses. Chez ces trois espèces, les glandes accessoires sont des plus petites, et ce n'est pas sans difficulté qu'on peut les observer dans le dernier segment du corps. Elles sont formées d'une houppe blanche et arrondie de vésicules microscopiques, inégales et pédiculées.
2. Organes mâles
On peut répartir les Chilopodes en deux groupes sous le rapport des organes reproducteurs mâles. Le premier groupe comprend les Lithobies et les Scutigères ; le second embrasse les Scolopendres, les Cryptops et les Géophiles.
PREMIER GROUPE.
Lithobius forcipatus (fig. 14).

Fig 14
Organes génitaux mâles du Lithobius forcipatus.
Cette espèce a déjà exercé le savant scalpel de M. Léon Dufour. MM. Treviranus et Stein ont également fait connaître l'appareil reproducteur mâle de ce Chilopode ; aussi passerai-je rapidement sur la description de ses organes génitaux.
Testicule.—La glande spermagène forme un tube impair, plusieurs fois replié, opaloïde et rigide. Son extrémité supérieure est très déliée, et fixée par un ligament suspenseur d'une grande ténuité ; son extrémité inférieure est également capillaire, mais sa région moyenne se renfle graduellement. Ce tube est la seule partie de l'appareil où j'aie observé les cellules mères des spermatozoïdes ; aussi lui donnerai-je le nom de testicule. M. Treviranus l'a également pris pour l'organe spermagène ; il le désigne sous le nom de vaisseau séminal du centre. C'est aussi l'opinion de M. Stein. M. Léon Dufour a vu des vésicules séminales dans ce boyau médian, et dans les deux tubes latéraux qui l'accompagnent.
Vésicules séminales.—Ces deux tubes sont cylindriques, partout d'égal diamètre et de moitié plus courts que le tube testiculaire. Dans leur état de turgescence séminale, ils sont d'un beau blanc de lait, et si gonflés de sperme, qu'à la moindre piqûre ce liquide s'écoule rapidement en jet d'apparence soyeuse. Pris dans ces tubes, le sperme se compose de pulviscule à grains très menus, et d'écheveaux de spermatozoïdes complètement développés. Mais on n'y trouve jamais les vésicules spermagènes observées dans le tube médian, et dans celui-ci on ne voit pas non plus la pulviscule qui gonfle les premiers. M. Stein n'a pas trouvé de spermatozoïdes dans les tubes latéraux ; je les ai vus quelquefois moi-même uniquement remplis de pulviscule ; mais le plus souvent ils contenaient, en outre, d'innombrables faisceaux de spermatozoïdes.
M. Treviranus les nomme réservoirs latéraux. M. Léon Dufour, qui les confond sous le même nom avec le tube testiculaire, les appelle vésicules séminales. M. Straus les prend pour deux testicules ; M. Stein leur réserve le nom d'épididyme. Parmi toutes ces dénominations, la plus convenable me paraît être celle de M. Léon Dufour. J'appellerai donc vésicules séminales les tubes latéraux des Lithobies. Leur fonction est de servir de réservoir au sperme, et de sécréter le liquide laiteux et la pulviscule qui se mélangent avec lui au sortir de la glande spermagène. Déférents.—Les deux vésicules confluent en arrière en une anse, dans la concavité de laquelle s'insère le tube testiculaire, tandis que de sa convexité partent deux canaux déférents, grêles et courts, formant eux-mêmes une autre anse qui embrasse le rectum, de manière que le tube digestif, placé au-dessous de l'appareil reproducteur dans la partie antérieure, passe au-dessus de lui dans les derniers segments, et que l'anus est supérieur à l'orifice génital. Les deux déférents débouchent dans un court renflement ovalaire qui termine l'appareil, et que M. Treviranus prend pour un pénis, mais qui ne constitue qu'un simple renflement éjaculateur.
Scutigera araneoides (fig. 15).
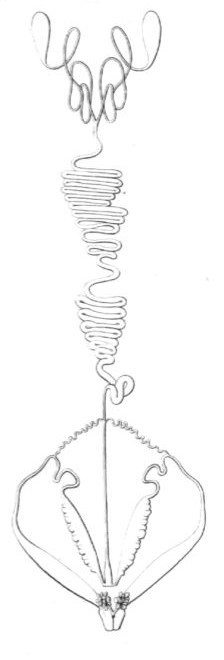
Fig 15
Organes génitaux mâles du Scutigera araneoides.
C'est encore M. Léon Dufour qui, dans ses infatigables investigations, nous a dévoilé la structure étrange des Scutigères ; aussi l'anatomie de ce Chilopode laisse-t-elle bien peu à désirer. J'ai été cependant assez heureux pour apercevoir quelques nouveaux détails d'organisation.
Testicules.—Ils sont au nombre de deux, et se composent de deux utricules ovoïdes, rigides, semi-diaphanes, et terminés chacun par un conduit capillaire, capricieusement entortillé, et se réunissant bientôt au conduit capillaire voisin pour former un tronc commun. Ce canal, d'un diamètre un peu plus considérable, se replie un grand nombre de fois, d'une manière fort élégante, de droite à gauche et de gauche à droite, et forme ainsi une sorte d'épididyme, qui recouvre d'une large bande la partie moyenne du tube digestif. En arrière, l'épididyme se divise en trois déférents : l'un médian rectiligne, les deux autres grêles et très flexueux. Le premier se rend dans un sinus profond, formé par la confluence des deux vésicules séminales ; les seconds relient le déférent moyen avec la partie supérieure du canal éjaculateur du même côté. Ces canaux latéraux ont échappé à M. Léon Dufour.
Vésicules séminales.—Ce sont deux sacs oblongs portant sur leur côté externe une vingtaine de petites vésicules sessiles qui rendent ce côté tout festonné. A l'issue de ces sacs commencent les deux conduits éjaculateurs qui se renflent peu à peu, et finissent par acquérir, dans leur état de turgescence, un diamètre considérable. Leur portion renflée est remarquable par son élasticité, qui fait jaillir violemment le liquide laiteux dont elle est gonflée, lorsque avec la pointe d'une aiguille on pique sa paroi.
M. Léon Dufour appelle testicules ces deux sacs festonnés, et il donne le nom de vésicules séminales aux deux utricules que je prends pour les glandes spermagènes. L'examen microscopique ne permet pas de se méprendre sur la nature de ces deux organes. A la fin de l'été les deux utricules terminaux renferment d'innombrables faisceaux de spermatozoïdes et des myriades de cellules spermatiques. Les deux conduits capillaires qui les continuent renferment aussi des spermatozoïdes, mais pas de cellules. Cette première partie de l'appareil reproducteur est opaloïde ; le reste, épididyme, vésicules séminales, conduits éjaculateurs, est d'un blanc de lait, et ne renferme pas encore de spermatozoïdes, mais un liquide tenant en suspension un nombre immense de corpuscules elliptiques, diaphanes et très menus. Parmi ces corpuscules s'en trouvent d'autres en plus petit nombre, régulièrement ovales, parfaitement diaphanes, et d'un diamètre une dizaine de fois plus grand. Entre ces deux extrêmes on voit d'autres corpuscules de grosseur intermédiaire, et qui démontrent qu'ils sont tous de même nature. C'est un spectacle admirable que ces myriades de petits corps : les uns, plus petits et plus nombreux, formant comme un fond de tableau tout semé de points diaphanes ; les autres, beaucoup plus grands, étalant çà et là leurs transparentes et larges ellipses. Les sacs festonnés doivent sécréter ce liquide laiteux et ces corpuscules elliptiques, car ils en contiennent encore plus que les autres organes. Les utricules terminaux, au contraire, n'en renferment pas la moindre trace, mais simplement des spermatozoïdes et leurs cellules mères. Je regarde donc ces derniers comme des testicules, et les sacs festonnés comme des vésicules séminales, comme les analogues des deux tubes latéraux des Lithobies, tubes qui sécrètent eux-mêmes un liquide laiteux tenant en suspension de menus corpuscules.
DEUXIÈME GROUPE.
Les autres Chilopodes présentent une uniformité remarquable dans leur appareil génital mâle. Les glandes spermagènes sont formées d'un nombre variable d'utricules fusiformes, tantôt isolés, tantôt réunis deux par deux, et toujours terminés aux deux extrémités par un conduit capillaire d'une grande ténuité, (lui se rend dans un canal médian commun également très délié.
Scolopendre complanata (fig. 16 [voir ci-dessus]).
Les utricules testiculaires, au nombre de 24, sont intimement accolés deux par deux, et forment ainsi 12 couples disposées en chaîne longitudinale, dans la première moitié des 19 derniers segments qui logent l'ensemble de l'appareil reproducteur. Chaque utricule produit à ses deux extrémités un canal capillaire assez court, qui s'unit étroitement avec le canal correspondant du second utricule de la même paire, mais sans se confondre avec lui. Enfin les deux canaux excréteurs d'une même extrémité de chaque couple plongent ensemble dans un canal longitudinal commun à toute la série. Ce canal commun se termine en avant par un bout délié, aveugle, qui sert de ligament suspenseur et s'attache sur le dos du jabot, entre les glandes salivaires. Son extrémité inférieure se continue avec l'épididyme.
Épididyme.—Je donne ce nom à un canal fort long, mais très étroit, d'un blanc jaunâtre, très sinueux, et entortillé un grand nombre de fois sur lui-même.
Bourse des spermatophores.—L'épididyme est suivi par un large boyau noduleux, irrégulièrement replié sur lui-même, où se forment les spermatophores, et qui sécrète l'enveloppe albumineuse de ces singuliers appareils fécondateurs.
Vésicule séminale.—A la bourse des spermatophores fait suite un canal droit ou conduit éjaculateur qui, à peu de distance de l'orifice génital, produit un boyau flasque, irrégulier, formant une anse dont les deux bouts communiquent à des hauteurs un peu inégales avec le conduit éjaculateur. J'ai fréquemment trouvé ce boyau plein d'une substance blanche formée de pulviscule à grains très menus, substance qui, dans la bourse aux spermatophores, se mélange avec le produit testiculaire. Cet organe me paraît être l'analogue dégénéré des tubes latéraux des Lithobies et des sacs festonnés des Scutigères. Je lui donne donc, comme aux premiers, le nom de vésicule séminale. Son rôle doit être fort secondaire, car on n'en trouve plus de traces chez les autres Scolopendrites. L'anse fermée que décrit cette vésicule séminale est traversée par le rectum.
Plusieurs auteurs se sont occupés de l'anatomie des Scolopendrites. Müller (6) et Kutorga (7) nomment Scolopendra morsitans l'espèce qui fait le sujet de leurs recherches. M. Straus (8) donne également quelques détails sur une Scolopendre à laquelle il applique le même nom. Comme le dit M. Lucas (9) on doit avoir confondu sous le même nom de Scolopendra morsitans des espèces différentes, car il serait impossible de se rendre compte autrement des résultats contradictoires où sont arrivés les anatomistes qui ont porté le scalpel dans les entrailles de ce roi des Chilopodes. Les deux tubes variqueux, reliés par des anastomoses, de la figure que donne J. Müller, ne rappellent en rien l'appareil testiculaire que je viens de décrire. La description que je trouve dans Straus concorde mieux avec ce que j'ai vu. Cet habile anatomiste ne décrit cependant que huit paires d'utricules testiculaires, tandis que j'en ai trouvé constamment douze paires. Enfin il décrit, dans la partie terminale de l'appareil, trois paires de glandes, dont deux ressemblent à deux petites masses de graisse, et dont une est filiforme. Je ne m'explique pas cette troisième paire, à moins que la vésicule séminale n'ait donné lieu à cette illusion. Pour éviter toute confusion, bien que le nom de morsitans soit appliqué par quelques auteurs à la Scolopendre du midi de la France, j'ai adopté la dénomination de Scolopendra complanata que Latreille lui donne (10). Sa description est d'ailleurs la seule que j'aie trouvée convenant parfaitement à l'animal que j'ai étudié, et qui abonde sur les collines boisées des environs d'Avignon.
Cryptops Savignyi et C. hortensis (fig. 17).
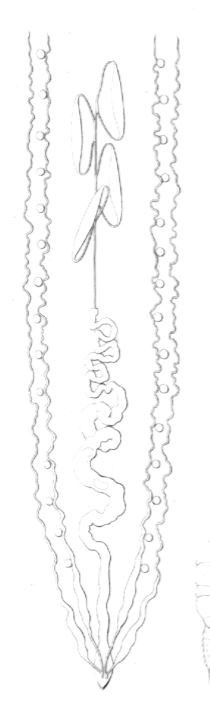
Fig 17
Organes génitaux mâles du Cryptops Savignyi.
L'appareil reproducteur mâle a la même conformation chez ces deux espèces congénères. Les testicules sont au nombre de quatre. Comme chez les Scolopendres, ils ont la forme d'utricules graduellement acuminés, et terminés par un conduit capillaire à chaque bout ; mais, au lieu d'être groupés par couples, ils sont indépendants l'un de l'autre. Le canal capillaire qui les relie se renfle brusquement vers l'origine du ventricule chylifique, et donne naissance à un conduit tortueux ou épididyme, suivi d'une portion encore plus large où se forment les spermatophores. Enfin l'appareil se termine par un canal éjaculateur droit et dépourvu de la vésicule séminale que possèdent les Scolopendres.
Geophilus (fig. 18).

Fig 18
Organes génitaux mâles du Geophilus Gabrielis.
Les quatre espèces que j'ai étudiées possèdent toutes deux utricules testiculaires fusiformes, reliés par les deux bouts à un canal capillaire commun.
L'épididyme est formé par un vaisseau délié, plié et replié dans tous les sens d'une manière inextricable. Lorsqu'il est développé, sa longueur est démesurée dans quelques espèces. Chez le G. Gabrielis, il atteint presque 1 décimètre, et chez le G. Ilicis il égale en longueur le corps de l'animal. Chez le G. convolvens, il est beaucoup moins long, capillaire à ses extrémités, et un peu renflé dans sa partie moyenne.
Après l'épididyme survient subitement un boyau beaucoup plus large et plus court, tantôt à peu près droit (G. convolvens), tantôt pelotonné. De ce boyau, part une arcade qui fournit deux conduits plus ou moins flexueux, de même diamètre et de même apparence que le canal unique d'où ils émanent ; c'est dans ces deux branches et dans la partie élargie du canal simple qui les précède que s'organisent les spermatophores. Les deux branches s'atténuent insensiblement en arrière, et se rejoignent de nouveau pour former un canal éjaculateur très court. Avant d'effectuer cette jonction, chacune se renfle, chez le G. Gabrielis, en une petite ampoule. Le rectum s'engage toujours dans l'anse allongée formée par cette partie terminale de l'appareil reproducteur.
Glandes accessoires.—M. Léon Dufour a vu quatre de ces glandes chez les Lithobies, mais il a laissé passer inaperçues celles des Scutigères ; Kutorga en a reconnu également quatre chez les Scolopendres ; Stein n'en attribue que deux aux Géophiles. D'après mes recherches, les Lithobies, Scolopendres, Cryptops et Géophiles, en ont quatre; les Scutigères n'en ont que deux.
Rien n'est plus varié que l'interprétation qu'on a donnée des glandes génitales postérieures des Chilopodes. J'ai déjà dit, en décrivant les organes femelles, que M. Treviranus les a prises pour des masses adipeuses ; M. Straus pour des organes urinaires. M. Léon Dufour les prend, chez les Lithobies mâles, pour les testicules ; M. Duvernoy (11) les compare aux prostates des Mammifères. Le mode de terminaison de leurs conduits excréteurs peut jeter quelque jour sur leurs fonctions problématiques, en démontrant que leur produit n'est pas déversé dans le canal vecteur du sperme, et ne peut être par conséquent assimilé soit au suc prostatique, soit à tout autre fluide destiné à se mélanger avec le fluide séminal. Chez les Chilopodes, l'anus et l'orifice génital sont renfermés ensemble dans un petit étui corné, composé de deux faibles écailles concaves placées l'une au-dessus, l'autre au-dessous. Cet étui gagnait-excrémentiel est rétractile, et l'animal peut le faire légèrement saillir ou l'abriter sous la dernière plaque dorsale. Sa cavité est divisée transversalement par une cloison charnue qui sépare les deux orifices. L'orifice supérieur est l'anus, l'autre est l'orifice génital. Chez la Scolopendre, l'écaille inférieure formant le plancher de l'orifice génital est revêtue à l'intérieur d'une lame charnue, linguiforme, et creusée d'une gouttière sur sa ligne médiane. De chaque côté de cette gouttière se voient, couchés contre les bords latéraux de l'écaille, deux petits tubes rigides et libres en arrière, et qui, en avant, plongent dans l'épaisseur des tissus. Leur extrémité libre arrive au même niveau que le bord postérieur de l'écaille qui les porte. En les suivant de proche en proche, on ne tarde pas à les voir se diviser chacun en deux canaux capillaires, qui sont les conduits excréteurs des deux glandes du même côté : l'une interne, plus petite ; l'autre externe, plus grande (fig. 16). Puisque leurs canaux sécréteurs se terminent à l'extrémité même de l'étui gagnait-excrémentiel, il est évident qu'elles ne fournissent pas un liquide complémentaire du sperme, destiné à se mélanger intimement avec lui. D'après une observation que je rapporterai bientôt, je crois qu'elles sécrètent une sorte de nidamentum, sur lequel le mâle dépose son produit, et, pour ainsi dire, pond ses spermatophores.
Lithobius forcipatus (fig. 14).
Les quatre glandes sont allongées, lancéolées, en forme, de sacs conoïdes, aplatis, et irrégulièrement vésiculeux à la surface. La paire externe est deux fois environ plus longue que l'autre ; ses déférents sont très courts. La paire interne a, au contraire, deux longs conduits excréteurs.
Scutigera araneoides (fig. 15).
Il n'y a ici que deux glandes ; elles sont fort petites, et composées de vésicules groupées à l'extrémité d'un pédicule assez gros, mais très court.
Scolopendra complanata (fig. 16).
Elles rappellent par leur forme celles des Lithobies.
Cryptops Savignyi et C. hortensis (fig. 17).
La paire interne a la forme remarquable des glandes des femelles, et se compose de deux longs vaisseaux sinueux portant latéralement une quinzaine de vésicules arrondies. La paire externe est formée de deux longs vaisseaux pareils aux premiers, mais dépourvus de vésicules.
Geophilus (fig. 18).
Les quatre espèces étudiées ont toutes deux paires de glandes. M. Stein n'en a vu qu'une paire chez le Geophilus subterraneus.
Les glandes internes sont environ trois fois plus courtes que les autres et flexueuses ; elles sont formées, les unes comme les autres, d'un sac cylindrique, très étroit relativement à sa longueur, et dont l'axe est parcouru par un canal diaphane, émettant de toutes parts, sur son trajet, des houppes glanduleuses, très touffues, en forme d'arbuscules. C'est ce canal qui, en se continuant hors du sac, forme le conduit excréteur.
Spermatophores.
Les spermatozoïdes des Chilopodes sont capillaires, et d'une longueur démesurée. Ceux des Géophiles sont contournés en pas de vis fort lâche, ce qui leur donne à plat une apparence régulièrement ondulée ; les autres n'ont jamais cet aspect. Chez les Lithobies et les Scutigères, les spermatozoïdes se groupent simplement en papillotes plus ou moins régulières, en écheveaux d'autant mieux formés qu'on les observe plus près de l'orifice génital. Mais chez les Scolopendres, les Cryptops et les Géophiles, ils sont renfermés dans des capsules communes d'une structure très remarquable, et analogues aux spermatophores des Céphalopodes et de quelques Crustacés.
Scolopendra et Cryptops.

Fig 19
Spermatophore du Scolopendra complanata.

Fig 20
Le même au moment de sa déhiscence.

Fig 21
Spermatophore du Cryptops Savignyi.
C'est dans la partie la plus renflée du canal vecteur du sperme, partie que j'ai appelée bourse aux spermatophores, que se forment ces appareils singuliers. Us spermatozoïdes traversent lentement l'épididyme, groupés en longs écheveaux. Arrivés dans cette bourse, ils se roulent en papillotes, qui se réunissent en grand nombre pour former un petit noyau blanc. Ce noyau se revêt d'abord d'une couche de pulviscule blanche, et enfin de deux tuniques superposées. Le nombre des spermatophores n'est pas considérable : je n'en ai jamais trouvé plus de sept, le plus souvent même moins. Complètement formés, ils ont 3 millimètres de diamètre chez la Scolopendre, et 1 millimètre chez les Cryptops. Leur forme est la même chez les deux genres, et se rapproche de celle d'un rein tricuspide (fig. 19, 20, 21) au côté concave. Leurs tuniques sont au nombre de deux. L'extérieure est très épaisse, transparente comme du cristal, et douée d'une grande élasticité qui la fait se tordre et se contourner même dans l'eau, quand on la découpe par lambeaux. Elle enveloppe étroitement de toutes parts le spermatophore, en laissant cependant au côté concave une boutonnière béante. A chaque coin de cette boutonnière, la tunique externe se renfle en petite dent cristalline. La seconde tunique est très fine, flasque, et forme une poche sans issue qui renferme des myriades de papillotes spermatiques. Par la boutonnière ou orifice de la première tunique, le sac interne fait hernie, et laisse échapper un mamelon d'un blanc mat, placé entre les deux dents cristallines (fig. 19-21). Dans l'eau, ce mamelon se gonfle, devient de plus en plus saillant, et finit par crever en vomissant un torrent de faisceaux de Spermatozoïdes convolutés. En un instant, le spermatophore est vidé par la contraction de la membrane élastique (fig. 20).
Géophilus (fig. 22).

Fig 22
Cylindres spermatiques du Geophilus convolvens dépouillés de leur enveloppe pulvérulente.
Les spermatozoïdes, au lieu de se convoluter en papillotes, s'enroulent plusieurs ensemble circulairement, et superposent leurs divers tours de spire, de manière à figurer un anneau, ou mieux un court cylindre creux de cordes enroulées. Chaque cylindre se revêt alors d'une couche protectrice, formée de pulviscule hyaline, faiblement agglutinée. Au moindre contact, cette enveloppe fugace se dissipe en légers flocons. On trouve ces cylindres par milliers dans les deux tubes parallèles terminant l'appareil reproducteur, et dans la portion renflée du canal qui les précède. Dans l'épididyme et plus haut, les spermatozoïdes sont simplement groupés en faisceaux. A l'oeil nu, ces cylindres apparaissent comme de petits points blancs, arrondis, à contour très net ; avec un peu d'attention, on peut même reconnaître que le centre de ces points circulaires est évidé. Mais ce n'est qu'avec le secours du microscope qu'on peut contempler tout ce qu'a de merveilleux cette admirable architecture spermatique ; alors, en effet, chaque point devient une couronne, un anneau ou plutôt un cylindre, rappelant on ne peut mieux la forme d'un tas de câbles empilés. Bientôt le spectacle s'anime ; une mystérieuse impulsion est communiquée à ces étonnantes machines, et chaque cylindre se met à tourner rapidement autour de son axe. Peu à peu les extrémités caudales des spermatozoïdes qui le composent se dégagent du groupe, s'échappent tangentiellement, et rayonnent en plus en moins grand nombre dans toutes les directions et, pendant que le cylindre tourne, chaque spermatozoïde enroule autour de lui sa partie libre, tout en exécutant des mouvements de reptation onduleux comme ceux d'un serpent. D'autres fois le cylindre se divise en deux, trois parties reliées par un filament ; alors la gyration de deux fragments voisins se fait en sens contraire, l'un s'enroule, l'autre se déroule, et le filament qui les relie accroît le premier aux dépens du second. Ces mouvements durent près d'une demi-heure avec la même vivacité, puis se ralentissent graduellement ; alors la gyration, au lieu de s'effectuer toujours dans un même sens, devient alternative, et à périodes d'autant plus courtes que le phénomène touche plus à sa fin. Un même cylindre s'enroule ou se déroule tour à tour, et les filaments qui s'en échappent tangentiellement s'en rapprochent ou s'en éloignent. Enfin tout rentre dans le repos, sans que le cylindre soit déformé. Mais il n'en est pas toujours ainsi : au moment de la plus grande violence de la gyration, les spermatozoïdes font effort pour se dégager, desserrent leurs tours de spire, et émettent loin du cylindre des boucles, dont les deux branches se tordent ensemble, autant que le permet leur longueur, puis se détordent pour recommencer immédiatement une nouvelle torsion. Lorsque ces boucles deviennent abondantes, le cylindre se détruit peu à peu ; les divers filaments spermatiques s'enlacent, s'enchevêtrent d'une manière inextricable, et avec tous ces mouvements discordants le désordre est bientôt au comble ; dans ce cas, la fin du mouvement s'annonce par de lentes oscillations, que les boucles exécutent en se tordant faiblement dans un sens, puis dans l'autre.
Enfin, disons-le, ce spectacle étrange, ces cylindres qui tournent sur eux-mêmes, qui s'enroulent ou se dévident, ces spermatozoïdes qui progressent en serpentant, tout cela n'est qu'une illusion, mais une illusion si parfaite, que ce n'est qu'en tenant des heures entières le regard attentivement fixé sur le microscope, que j'ai pu me rendre compte de ces bizarres apparences. En examinant l'extrémité caudale d'un spermatozoïde qui paraît s'enrouler sur un cylindre, on reconnaît que, malgré l'apparence d'une vive progression du filament, son point terminal n'avance pas, et fouette constamment les mêmes corpuscules en suspension dans le liquide. En outre, un examen soutenu de cette extrémité démontre que le filament spermatique n'a pas la forme d'une ligne ondulée située dans un même plan, comme le font croire les premières apparences, mais bien celle d'une hélice à tours de spire largement distants. Cette forme hélicoïde explique parfaitement l'illusion produite. Qui ne sait, en effet, qu'un tire-bouchon qu'on fait tourner autour de son axe paraît progresser dans un sens ou dans l'autre, suivant le sens de sa rotation, sans réellement changer de place. Les spermatozoïdes des Géophiles n'ont donc pas de mouvement de translation, mais simplement un mouvement révolutif autour de leur axe, mouvement qui, combiné avec la forme hélicoïde, produit la gyration illusoire des cylindres, et la reptation apparente des filaments spermatiques.
Pourquoi tout ce luxe de précautions au sujet de l'élément fécondant ? Pourquoi ces spermatophores, ces capsules hermétiquement closes, qui ne doivent éclater qu'à un moment donné ; ces glandes génitales postérieures, si constantes dans leur nombre, si variables dans leur forme, mais toujours remarquables par leur grand développement ? Pourquoi ces grappes sécrétoires, ces sacs glandulaires, ces cordons de vésicules, qui attendent le sperme à son passage pour le revêtir, sans doute, d'une nouvelle enveloppe protectrice ? L'élément mâle n'est donc pas introduit directement dans l'organisme de la femelle, puisqu'il est si soigneusement prémuni contre les agents extérieurs ; en d'autres termes, il n'y a donc pas d'accouplement ?
Aucun Chilopode ne possède d'organe copulateur, car on ne peut donner, comme le fait M. Treviranus, le nom de pénis au léger renflement qui termine le canal vecteur séminal des Lithobies. Si l'accouplement a lieu, il ne peut s'opérer que par le retournement de la portion terminale du conduit éjaculateur. D'un autre côté, personne, que je sache, n'a été témoin de cet accouplement. Je n'ai pas été plus heureux. Deux années entières j'ai poursuivi mes recherches sur ces animaux; jamais je n'en ai vu d'accouplés, pas même parmi les nombreux individus de tout genre que je tenais en captivité, et que j'avais sans cesse sous les yeux.
Ce n'était pas, du reste, sans difficulté que je parvenais à maintenir dans chaque vase un nombre de mâles en rapport avec celui des femelles. La cohabitation des deux sexes donnait invariablement lieu à d'atroces tragédies, car les femelles, plus vigoureuses, rongeaient impitoyablement la tête de leurs mâles. Les Scolopendres surtout montrent à un haut degré cette étrange antipathie trahissant des amours sanglantes que le mâle ne saurait satisfaire sans encourir de graves périls pour sa vie. En liberté, cette antipathie s'étend même entre des individus de même sexe. Il ne m'est pas arrivé une seule fois de rencontrer deux Scolopendres au même gîte, si jeunes qu'elles fussent. On sait que chez les Aranéides, les amours sont tout aussi tragiques, ce qui n'empêche pas le mâle de fléchir sa femelle et d'opérer la fécondation ; aussi je me hâte d'arriver à un fait plus concluant.

Fig 23
Spermatophore du Geophilus convolvens déposé sur son réseau aranéeux.
A la fin de septembre, en examinant des Geophilus convolvens, que je tenais depuis quelque temps en captivité, j'aperçus dans les couloirs pratiqués par ces animaux, au sein du terreau qui leur servait de demeure, de très petits réseaux formés de filaments aranéeux et disposés de loin en loin (fig. 23). Leur trame se composait d'un petit nombre de fils tendus en travers d'une paroi à l'autre du couloir, et irrégulièrement croisés. D'autres réseaux pareils se montraient hors du terreau, et étaient tendus entre quelques brins de mousse dont j'avais achevé de remplir le bocal. Au centre de chacun était appendu, loin du contact de tout corps étranger, un globule sphérique blanc de la grosseur d'une petite tête d'épingle, et que je pris d'abord pour un oeuf. Ayant détaché un de ces réseaux, je soumis au microscope le globule dont l'enveloppe délicate éclata au premier contact. Quel ne fut pas mon étonnement en reconnaissant dans ce globule une gouttelette de sperme avec ses cylindres de spermatozoïdes en pleine giration ! Plusieurs autres furent recueillis, les résultats firent les mêmes. Quelque inexplicable que fût ce fait si exceptionnel, le doute était impossible ; j'avais bien sous les yeux du sperme de Géophile. Ait bout de deux ou trois jours, ces premiers globules avaient disparu, soit desséchés, soit ravagés par des Acarus, qui s'en montraient friands. Mais de nouveaux réseaux étaient déjà tendus avec leur gouttelettes au centre, et ainsi de suite pendant un mois et demi environ ; de sorte que malgré leur prompte destruction, il y en avait constamment cinq ou six dans le flacon. J'observai les derniers le 12 novembre.
Ce fait s'est donc répété trop souvent et avec trop de régularité pour pouvoir être regardé comme accidentel. Ainsi il est démontré expérimentalement que les Géophiles, et par suite les autres Chilopodes, déposent leurs spermatophores sur un réseau aranéeux, sur un nidamentum qui les isole et les protège du contact des corps étrangers qui pourraient les souiller. Le fluide séminal devant ainsi rester quelque temps livré à l'action des agents extérieurs, on se rend compte de ces glandes génitales postérieures dont on ne savait que faire, et qui doivent servir à tisser le nidamentum ; on s'explique enfin la nécessité de ces capsules protectrices du sperme si solides, si soigneusement closes chez les Scolopendres et chez les Cryptops, capsules d'ailleurs contradictoires avec un accouplement.
Que devient ce spermatophore ? L'observation m'a fait défaut au moment où, après de nombreuses tentatives infructueuses, je commençais à espérer la révélation de ce mystère physiologique. Bien qu'il y eût des femelles dans le même vase, aucune n'a répondu à l'appel du mâle, car je n'ai pas obtenu de ponte. A en juger par leur taille, et par leurs ovaires que j'ai examinés quand les mâles ont cessé de filer leurs réseaux spermatiques, j'ai lieu de croire qu'elles n'avaient pas encore atteint l'âge nécessaire à la maturité des ovules.
On pourrait croire que la femelle vient déposer ses oeufs autour du spermatophore qui, ramolli au moyen des humeurs sécrétées par les glandes génitales, éclate et chasse son contenu. La fécondation s'opérerait alors par l'infiltration du sperme à travers le tas d'ovules. Mais alors d'où proviendrait le sperme des réservoirs séminaux des femelles ? Faut-il, avec M. Stein, admettre, qu'il se forme dans ces appareils ? L'observation m'a prouvé le contraire. Je crois plutôt qu'avant la ponte, la femelle en possession d'un spermatophore le fait éclater, soit sur le réseau nidamentaire, soit dans sa vulve, en l'arrosant du produit de ses glandes génitale, et en conserve le contenu dans ses réservoirs jusqu'à l'expulsion des ovules, qui seraient fécondés au moment de leur passage devant les mamelons où se rendent les déférents de ces réservoirs. Lesspermatozoïdes, que ces derniers contiennent dans toutes les saisons, proviennent ainsi du fluide séminal non employé. Leur rigidité, leur manque de mouvement démontrent qu'ils ne forment plus qu'un résidu désormais impropre à la fécondation : c'est ce que l'expérience confirme pleinement. En juin, un Geophilus electricus pondit même sous mes yeux. Les oeufs au nombre d'une trentaine, étaient ronds. blanchâtres et groupés en petits tas. La ponte achevée, la mère s'est roulée en spirale autour de ses oeufs, qu'elle paraissait surveiller ; mais quelques heures après, je l'ai surprise à les dévorer avec avidité. Je suis cependant parvenu à en sauver la majeure partie que j'ai mise en sûreté, et j'ai soumis au scalpel les entrailles de la mère dénaturée. J'ai constaté que les vésicules séminales renfermaient des spermatozoïdes, aussi ai-je compté sur la prochaine éclosion de ces oeufs précieux. Vain espoir ! en peu de temps ils se sont moisis et complètement détruits. Ils n'étaient donc pas fécondés, et la mère n'a pas été aussi dénaturée que je le croyais : elle n'a fait que détruire une ponte stérile. J'ajouterai que, dans le vase, qui renfermait cette femelle, je n'avais jamais observé de réseaux spermatiques. C'est là, sans doute, la cause du peu de succès de celle ponte.
Quelle que soit la manière dont s'effectue la fécondation, il me paraît établi qu'elle se fait sans accouplement. L'absence complète d'organes copulateurs, les capsules protectrices du sperme, l'antipathie de ces hideux animaux, les réseaux spermatiques des Géophiles, tout me fait croire que le mâle dépose furtivement les spermatophores sur des réseaux tendus dans les couloirs souterrains ; et que c'est là que la femelle, guidée par son instinct et pressée par son fardeau, doit venir trouver l'élément complémentaire de ses ovules.
CHAPITRE II
DÉVELOPPEMENT
Lithobius forcipatus.
M. P. Gervais a fait connaître en partie l'évolution de cette espèce (12). Mes observations confirment et complètent celles du savant professeur.
Ayant recueilli, dans un Saule à demi pourri, du terreau où j'avais vu beaucoup de Lithobies, je fus assez heureux pour y trouver des jeunes en abondance et même des oeufs qu'il me fallut rechercher un à un. Ces oeufs sont ronds, d'un blanc jaunâtre, et de 1 millimètre environ de diamètre. En ayant ouvert un, je pus en extraire une jeune Lithobie possédant six paires de pattes, et les rudiments d'une septième paire sous forme de deux petites pointes coniques. Ainsi, dans l'oeuf même, l'évolution des pattes n'est pas simultanée, et s'opère d'avant en arrière, comme après l'éclosion. Les oeufs recueillis ne tardèrent pas à éclore, et, le 1er mai, je pu observer quelques Lithobies qui en provenaient.
1er stade.—Au sortir de l'oeuf, le jeune est composé de 10 segments, non compris la tête, et possède 4 paires de pattes et 7 articles aux antennes. Sa couleur est d'un beau blanc, excepté deux petits points rougeâtres placé de chaque côté de la tête, et qui sont les deux premiers ocelles. Sa longueur atteint 2 millimètres au plus. Sur l'avant-dernier segment apparaissent bientôt deux tubercules, qui s'allongent peu à peu en s'accolant aux côtés de l'anneau anal, dont ils atteignent l'extrémité. Ces deux pointes coniques deviennent enfin des pattes.
2ème stade. — 11 segments; 8 paires de pattes ; 11 articles aux antennes ; 2 ocelles rougeâtres de chaque côté. La couleur a changé. La tête est légèrement ambrée, et le reste du corps faiblement violacé. Deux paires de pattes rudimentaires apparaissent ensuite en arrière, et le nombre des articles antennaires augmente ainsi que celui des ocelles.
3ème stade. — 13 segments : 10 paires de pattes ; 17 articles aux antennes ; 4 ocelles de chaque côté, dont deux plus grands en arrière de la série. Deux nouvelles paires de pattes se forment bientôt à la suite des autres, et l'animal arrive au stade suivant.
4ème stade.— 15 segments : 10paires de pattes ; 22 articles aux antennes ; 4 ocelles de chaque côté. En peu de jours, sous la quatorzième plaque dorsale, se montrent simultanément 3 paires de pattes rudimentaires. Au stade suivant, le nombre des pattes doit donc être compté.
5ème stade. — 17 segments; 15 paires de pattes ; 26 articles aux antennes; 6 ocelles de chaque côté sur deux rangées, et d'autant plus petits qu'ils sont plus antérieurs. Longueur, 6 millimètres.
C'est à la fin de juin que j'ai observé ce dernier degré de développement. Les Lithobies mettent donc à peu près deux mois pour acquérir les huit paires de pattes et les sept anneaux qui leur manquent au sortir de l'oeuf. Les ocelles et les articles antennaires continuent encore longtemps à augmenter en nombre, sans qu'il y ait même de limite bien précise à celle augmentation.
Scutigera araneoides.
Je n'ai pas été témoin de l'éclosion des oeufs de Scutigère ; mais en recueillant, au mois d'août, des jeunes plus ou moins avancés dans leur évolution, et en les élevant, j'ai reconnu chez eux cinq stades comme chez les Lithobies, stades caractérisés par 7-9-11-13 et 15 paires de pattes. Les individus munis seulement de 7 paires de pattes se rapportent bien certainement à l'éclosion. Leur couleur pâle faiblement violacée, leur longueur de 2 millimètres à 2 millimètres 1/2 ne permettent pas d'en douter. D'ailleurs la conformité remarquable d'organisation des Scutigères et des Lithobies, leur mode commun de développement, démontrent à priori que le premier stade des Scutigères doit être le même que chez les Lithobies. Ainsi l'évolution des Scutigères offre une grande ressemblance avec celle des Lithobies ; mais elle est plus régulière, car l'accroissement du nombre de pattes est constamment de deux paires d'un stade à l'autre, tandis que, chez les Lithobies, il est d'abord d'une paire, puis à plusieurs reprises de deux paires, et enfin de trois. Le nombre des articles des antennes et celui des ocelles participent à cet accroissement. Dans les individus les plus jeunes, j'ai compté de 60 à 70 ocelles de chaque côté. J'en ai reconnu environ 150 chez un adulte. Enfin les articles des tarses croissent également en nombre ; les jeunes, possédant 7-9-11 paires de pattes, ont seulement de 20 à 22 articles aux tarses. Dans un adulte, j'ai obtenu les nombres suivants, qui présentent ce fait remarquable d'être sensiblement égaux à égale distance des extrêmes d'une série, dont le premier terme est le nombre d'articles des antennes, et le dernier le nombre d'articles des tarses postérieurs :
| Antennes | 400 art. | 15e tarse | 180 art. |
| 1er tarse | 49 | 14e | 47 |
| 2e | 43 | 13e | 43 |
| 3e | 41 | 12e | 41 |
| 4e | 40 | 11e | 39 |
| 5e | 40 | 10e | 39 |
| 6e | 38 | 7e | 37 |
| 9e | 36 | 8e | 36 |
Il n'est pas moins remarquable que la première moitié de la série embrasse l'ensemble des organes appendiculaires du jeune au sortir de l'oeuf, et la seconde moitié les appendices qui proviennent du bourgeonnement. Dans la première moitié du corps sont contenus les organes des sens et les parties essentielles de l'appareil digestif, en un mot ce qui est nécessaire à la conservation de l'individu ; dans l'autre moitié est logé l'appareil qui a rapport à la conservation de l'espèce. Ne dirait-on pas qu'à partir de sa région médiane l'animal se répète, mais en sens inverse ; qu'à la suite de l'individu, tel qu'il sort de l'oeuf, et capable de se suffire à lui-même, mais impuissant pour l'espèce, il s'en forme par bourgeonnement un autre chargé de la propagation ? Une profonde modification de l'écusson dorsal de la région médiane marque la séparation de la partie antérieure d'origine ovulaire, et de la partie postérieure d'origine gemmaire. En effet, le dernier écusson du jeune, au sortir de l'oeuf est précisément le plus grand de tous citez l'adulte, et si différent des autres que Linné l'a appelé élytre (13). Cet écusson recouvre trois paires de pattes, dont les deux premières appartiennent au jeune dès l'éclosion, et la troisième à la partie du corps produite par bourgeonnement. Il est frappant de retrouver une modification analogue chez les Lithobies. Chez ces Chilopodes, les écussons dorsaux sont alternativement étroits et élargis, à partir de celui qui recouvre les pinces. Celle loi se maintient parfaitement jusqu'au huitième qui est large ; mais le neuvième, qui devrait être étroit, est au contraire large comme le huitième ; par delà, la loi d'alternance reparaît sans nouvelle interruption. Il y a donc après le huitième écusson une interversion dans la loi qui préside à la première moitié ; or ce huitième anneau recouvre précisément la septième paire de pattes, c'est-à-dire qu'il est le dernier pédigère de ceux que possède le jeune lors de l'éclosion.
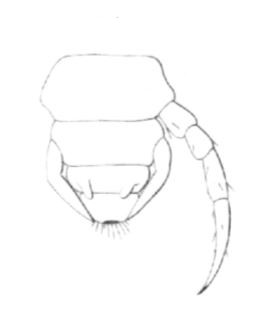
Fig 24
Derniers segments d'un jeune Lithobie forcipatus, à un degré d'évolution intermédiaire entre le 1er stade et Ie 2e.
La dernière paire de pattes complètement développées est la 7e paire, et est portée par le 8e segment. La 8e paire, qui doit être parachevés au stade suivant, atteint déjà l'extrémité anale. La 9e paire commence à poindre sous forme de deux très petits mamelons.
L'évolution produit chez les Insectes deux organismes successifs et complémentaires : la larve qui vit pour l'individu, l'insecte parfait qui vit pour l'espèce ; organismes qui passent de l'un à l'autre par les mystérieuses transformations de la nymphose. Il me semble voir un intime rapport entre la partie d'origine ovulaire des Chilopodes et la larve, entre la partie d'origine gemmaire et l'insecte parfait. Mais ici la larve n'est pas transformée de fond en comble pour atteindre l'état parfait ; elle est continuée par l'organisme reproducteur; il n'y a pas métamorphose d'un organisme relatif à la conservation de l'individu en un autre ayant rapport à la procréation de l'espèce, il y a simple gemmation du second par le premier. A ce point de vue, les modifications que l'âge amène chez les Chilopodes ne méritent pas le nom de métamorphose. Ces traces de dualité qui se manifestent dans la structure des Lithobies et des Scutigères ne rappellent-elles pas aussi d'une manière frappante ce qui se passe chez les Syllis, chez les Myrianides ? Supposons que, chez ces Annélides, l'organisme, l'individu reproducteur produit par bourgeonnement, soit unique, et ne se détache pas de la souche mère, n'aurons-nous pas à peu près la structure de ces deux Chilopodes ? Mais je m hâte de quitter ces considérations, qui, n'étant pas encore suffisamment mûries, laissent un champ trop libre à l'imagination.
Cryptops hortensis.
Dans un vase où je tenais des Cryptops hortensis, j'aperçus, au commencement de juillet, des jeunes qu'il m'est impossible de rapporter à un autre Chilopode que le précédent, bien que M. Paul Gervais en ait fait un genre nouveau sous le nom de Scolopendrella, genre qu'il place dans sa famille des Géophilides (14). N'ayant pas été témoin de l'éclosion, j'ignore si j'ai réellement observé le premier stade de l'évolution.
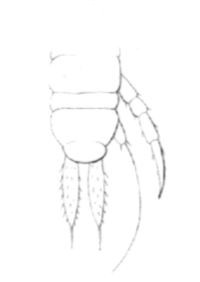
Fig 25
Derniers segments d'un jeune Cryptops hortensis muni de 8 paires de pattes parachevées. L'avant-dernier segment porte deux mamelons cirrifères, rudiments des pattes de la 9e paire. Les appendices du dernier segment sont les rudiments des grosses pattes terminales de l'adulte.
Les plus petits ont 1 millimètre 1/2 de longueur, huit paires de pattes et treize articles aux antennes. Le dernier segment (fig. 25) porte deux appendices exactement terminaux, rangés côte-à-côte parallèlement à l'axe du corps, et dirigés en arrière. Ils sont formés d'un seul article lancéolé, tout hérissé de cils, et portant au sommet un piquant roide , aigu et fort long. Ces appendices sont évidemment les rudiments des deux longues et grosses pattes qui terminent le corps des Cryptops, pattes que l'animal projette en arrière, sans les employer pour la marche. L'avant-dernier segment porte de chaque côté un petit mamelon conoïde, couronné par quelques cils, et muni au sommet d'un cirre long et roide, courbé en arc, et dépassant la plus reculée des appendices terminaux. Ce cirre exécute des mouvements trépidatoires, des oscillations plus ou moins rapides, et pourrait bien être, de quelque usage pour la locomotion. Les deux mamelons cirrifères forment les rudiments d'une nouvelle paire de pattes. Tous les autres segments sont pédigères, et ne présentent rien de particulier.
Ce que la tête présente de plus remarquable, c'est l'absence des diverses pièces de la bouche. Je suis simplement parvenu au microscope à apercevoir, à l'extrémité de la tête, un ostiole arrondi, un peu rembruni, et cilié sur les bords. Par la compression, j'ai mis à nu deux tubercules hyalins, très courts, faiblement recourbés, et placés de chaque côté de l'ostiole. Ces deux pointes obtuses pourraient bien être les rudiments des forcipules.
M. Paul Gervais signale deux petits stemmates sur la tête de sa Scolopendrella. J'ai, en effet, trouvé en arrière de la base des antennes deux points ronds, très bien circonscrits, mais qu'il m'est impossible de prendre pour des yeux. A la lumière réfléchie, ces points sont d'un blanc mat, et se détachent nettement sur le fond hyalin de la tête. Vus par réfraction, ils deviennent sombres, presque noirs. On ne peut expliquer ces apparences qu'en admettant que les deux prétendus stemmates sont simplement deux points où les téguments de la tête ont une plus grande opacité qu'ailleurs ; au reste, ces deux points ne se montrent quelquefois ni par réflexion, ni par réfraction.
Depuis le mois de juillet jusqu'au mois de décembre, j'ai vu mes jeunes Cryptops successivement avec 8, 9, 10 et 11 paires de pattes. Les articles des antennes étaient en même temps au nombre de 13, 14, 14 ou 15, 14 à 19. Muni de 11 paires de pattes, l'animal a 3 millimètres longueur. Sa couleur est encore d'un beau blanc.
Cryptops Savignyi.
Je rapporte à cette espèce de jeunes Chilopodes, qui offrent la plus grande ressemblance avec les jeunes du Cryptops hortensis. Je les ai recueillis sous les pierres des collines boisées des environs. Ils différent des premiers par leur plus grande taille, leurs antennes plus longues, et surtout par leurs plaques dorsales profondément échancrées en arrière et bordées d'un liséré d'un blanc mat. Ils ont de même les deux taches pseudo-oculaires, les deux pattes rudimentaires terminales et les deux mamelons cirrifères. Les pièces de la bouche leur manquent également. Je crois pouvoir les rapporter au Scolopendrella notacantha de M. Gervais (15).
En juin, un jeune de 1 millimètre 1/2 de longueur m'a montré 8 paires de pattes, les deux tubercules cirrhifères, les appendices terminaux et enfin 13 articles aux antennes. Ce résultat concorde en tout point avec ce que j'ai observé chez les jeunes du Cryptops hortensis. De cette époque jusqu'en décembre, j'ai pu recueillir des jeunes de 1 millimètre 1/2 à 4 millimètres de longueur et portant de 8 à 12 paires de pattes. Enfin j'ai vu les antennes acquérir dans cette période jusqu'à 28-29 articles. Les Cryptops adultes n'ont que 17 articles aux antennes. Nous avons donc ici un fait inverse de tous les précédents, c'est-à-dire un plus grand nombre d'articles antennaires chez les jeunes que chez les adultes. Il faut donc qu'avec I'âge ce nombre diminue, soit par la soudure, soit par la chute de quelques articles.
Après avoir acquis 11-12 paires de pattes, les jeunes, tant du Cryptops hortensis que du Cryptops Savignyi, entrent dans une longue période stationnaire dont j'ignore encore l'issue. J'ai tenté d'en élever dans un flacon rempli de terre. La plupart ont péri ; cependant , après plus de six mois de captivité, il m'en reste encore quelques-uns, et pendant ce long espace de temps, leurs dimensions, leur couleur, ainsi que le nombre de leurs pattes et de leurs articles antennaires, n'ont pas varié. Cette lenteur de l'évolution des Cryptops est d'autant plus frappante que nous avons vu les Lithobies et les Scutigères atteindre rapidement, au contraire, la forme adulte.
Scolopendra complanata.
J'ai vainement tenté fort longtemps d'élever cette espèce pour suivre son évolution ; le massacre des mâles par les femelles a rendu infructueuses toutes mes tentatives, et je n'ai pu obtenir de ponte. Je n'ai pas été plus heureux en recherchant des jeunes ; la plus petite Scolopendre que j'aie vue avait 24 millimètres de longueur, c'est-à-dire environ le cinquième de la longueur de l'adulte ; elle était d'ailleurs absolument conformée, sous tous les rapports, comme les individus ayant leur entière croissance. Mais il y a loin de cette taille de 24 millimètres à celle que le jeune doit avoir à l'éclosion ; aussi la parfaite ressemblance, tant pour l'organisation interne que pour l'organisation externe, des Crytops et des Scolopendres ne permet pas de douter que ces dernières ne subissent avec l'âge des modifications aussi profondes que les Cryptops. M. P. Gervais a cependant observé une femelle de Scolopendre qui, placée encore vivante dans un flacon d'alcool, y pondit non des oeufs, mais des petits déjà développés, ci ayant leur nombre normal de pattes et d'anneaux. L'espèce qui a présenté ce fait si exceptionnel d'ovoviviparité doit considérablement différer dans son organisation de celle qui m'a occupé, et qui, ne possédant aucun organe d'incubation interne, est certainement ovipare.
Geophilus.
Je ne connais pas non plus les premières phases de l'évolution des Géophiles. J'ai pu cependant constater que le nombre de leurs anneaux et de leurs pattes croît avec l'âge. Un jeune Geophilus Ilicis de 6 millimètres de longueur possédant seulement 37 paires de pattes ; la même espèce adulte en a jusqu'à 97 paires. Les autres espèces m'ont fourni des résultats analogues. J'ai constaté, en outre, chez le Geophilus electricus, que les nouvelles paires de pattes apparaissent une à une comme chez les Cryptops, et non plusieurs paires ensemble comme chez les Lithobies et les Scutigères. La nouvelle paire se développe sur l'avant-dernier segment, et apparaît d'abord sous forme de deux mamelons pareils à ceux des Cryptops, mais dépourvus de cirrhe. Quant aux artistes antennaires, je les ai toujours vus au nombre de 14, même dans celui qui n'avait que 6 millimètres de longueur.
Il était admis, comme règle générale, que les Myriapodes naissent avec trois paires de pattes. Cette règle paraît être sans exception pour les Chilognathes ; mais elle ne s'applique nullement aux Chilopodes. Les observations précédentes, quoique très incomplètes encore, démontrent que les Lithobies et les Scutigères naissent avec sept paires de pattes ; les Cryptops, et par suite les Scolopendres, très probablement avec huit paires ; et les Géophiles, suivant toute apparence, avec un plus grand nombre.
Le doyen de la Faculté des sciences,
Note
- Ann. des sc. nat., 1ère série, t. II.
Retour
- Anat. comp., t Il.
Retour
- Ann. des sc. nat., t. II, 1834.
Retour
- Anat. comp., p. 480.
Retour
- Ne pouvant rapporter ces deux Géophiles à aucune des espèces décrites par les auteurs, j'en donnerai ici les caractères:
1. Geophilus Ilicis. Tête ferrugineuse, cachant en entier les forcipules. Antennes pâles, de deux à trois fois aussi longues que la tête, composées de 14 articles diminuant graduellement de longueur de la base au sommet. Corps d'un jaune-paille, très pâle dans la région postérieure. Mâle : 96 paires de pattes ; appendices postérieurs sans onglet. Longueur, 80 millimètres. Femelle : 72 paires de pattes ; appendices postérieurs avec onglet. Longueur, 70 millimètres.
Dans les deux soies les appendices postérieurs sont conformés comme les pattes ordinaires, mais plus longs et composés de 6 articles, tandis que les pattes n'en ont que 5.—Commun autour des souches d'yeuse, et sous les pierres des collines boisées des environs.
2. Geophilus convolvens. Tête très petite, ferrugineuse, couvrant en entier les forcipules. Antennes deux fois environ plus longues que la tête, ferrugineuses, composées de 14 articles moniliformes. Corps graduellement acuminé aux deux extrémités, plus large au milieu. Dos brun avec une ligne étroite et médiane pâle. Les flancs et le ventre pâles. Mâle : 66 - 68 paires de pattes ; appendice postérieurs élargis en palette, ferrugineux, de 6 articles sans onglet. Longueur, 62 millimètres Femelle : 75 - 86 paires de pattes ; appendices postérieurs conformés comme les pattes ordinaires, de 6 articles avec onglet terminal. Longueur, 70 millimètres.
Reconnaissable au premier aspect par la manière dont il se pelotonne en boule en s'enroulant sur lui-même lorsqu'on l'irrite. Il diffère, en outre, des trois autres Géophiles mentionnés dans ce travail par ses deux glandes vénénifiques en forme d'utricules, dont le canaux excréteurs débouchent vers la pointe du forcipules. Ces glandes sont accompagnées de deux glandes salivaires, qu'on retrouve seules chez les trois autres espèces.—Commun dans les jardins.
Retour
- Zür anatomie der Scolopendra morsitans (lsis, 1829).
Retour
- Scelopendroe morsitantis anatome.
Retour
- Anat. comparative.
Retour
- Hist. mat. des Crustacés, des Arachnides et des Myriapodes.
Retour
- Nouveau Dict. d'hist. nat.
Retour
- Cuvier, Anat. comp., 2ème édit., t. VIII.
Retour
- Ann. des sc. nat ., 2ème série, t. VII.
Retour
- Systema naturae .
Retour
- Ann. des sc. nat., 3è série, t. II.
Retour
- Ann. des sc. nat., 3e série, t. II.
Retour